




Introduction
Célèbre pour ses figurations pariétales de l’Aurignacien et du Gravettien ancien particulièrement bien préservées, la grotte Chauvet Pont d’Arc (Ardèche) renferme également de nombreux indices des fréquentations humaines et animales sur les sols dans différentes galeries. Étudiée par Michel Alain Garcia (1944-2008), l’ichnologie constitue l’autre originalité singulière de cette cavité ornée, par la reconnaissance de quelques pieds et talons d’homme anatomiquement moderne, de nombreuses pistes d’ours des cavernes (Ursus spelaeus) ainsi que par des empreintes attribuées à des grands canidés1. Les documents ichnologiques ont souvent été signalés dans des gisements pléistocènes relatifs aux ursidés2 , félidés, canidés3 ou hyénidés4. Dans de nombreuses grottes ornées, des empreintes humaines ont été identifiées sur des sols paléolithiques (Le Tuc d’Audoubert5, Le Mas d’Azil6, Niaux7 en Ariège, Pech-Merle8 dans le Lot, Cussac9 en Dordogne, Aldène10 dans l’Hérault). Des fréquentations humaines associées à des canidés (chien/loup ?) ont récemment été mises en évidence pour la fin du Paléolithique supérieur, dans la grotte de la Basura en Italie11. Ces enregistrements des activités humaines et animales sont importants, dans une perspective géo-chronologique, paléoéthologique et anthropozoologique (occupations des différents espaces souterrains par les hommes et les prédateurs, relations hommes – carnivores).
Dans la grotte Chauvet Pont d’Arc, les études paléontologiques ont permis d’identifier une vingtaine de vestiges osseux attribuables à des grands canidés12. Des mesures ont directement été effectuées sur les ossements accessibles ou par photogrammétrie. Les empreintes pouvant être attribuées à ce groupe zoologique ont également été recherchées, localisées et, analysées avec le même protocole d’étude.
Le présent article dresse un bilan conjoint de la répartition spatiale des restes osseux et des empreintes appartenant aux genres Canis et Cuon et fournit des premiers éléments ostéométriques et ichnométriques pour le loup (Canis lupus) et le cuon (Cuon alpinus), à partir de données comparatives modernes et pléistocènes. L’implication anthropozoologique concernant l’identification spécifique des canidés étant importante pour le Paléolithique supérieur13 , il apparaît opportun d’essayer de déterminer dans un premier temps les données de la grotte Chauvet Pont d’Arc, avant d’envisager des occupations naturelles ou non de la cavité par ces espèces.
Matériel et méthodes
Conditionnées par les impératifs de conservation, les recherches à la grotte Chauvet Pont d’Arc s’effectuent à partir de cheminements balisés dans les différents secteurs aménagés d’une part et par des observations à distance (de visu, couverture photographique, lasergrammétrie, photogrammétrie) des sols sur lesquels restes osseux et empreintes sont visibles d’autre part. Toutes les observations sont effectuées in situ, aucun échantillon n’étant manipulé ou sorti de la cavité.
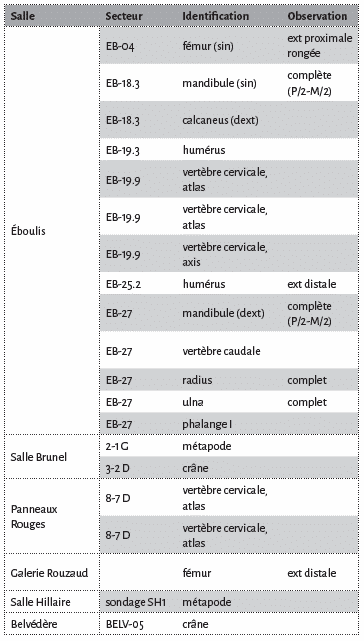
Le nombre de vestiges paléontologiques est de 20 (Tab. 1), correspondant à 4 individus (atlas complets). Des mesures ont été prises sur quelques pièces selon la nomenclature de von den Driesch14, assorties de données comparatives pour le loup15 et le cuon16 modernes et pléistocènes.
Pour l’ichnologie, les codes de mesurage des empreintes de loup proposés par Harris & Beam17, repris par Halfpenny18, ont été retenus, complétés par des mesures de Ledoux & Boudadi-Maligne19. Une correspondance entre ces méthodes est proposée (Fig. 1). Afin de disposer de spécimens directement utilisables, des mesures ont été prises sur des empreintes de loup de Pologne provenant de Bialowieza et de Bieszscady (Fosse & Selva, inédit) et des moulages d’empreintes de cuon captif ont été réalisés par le Zoo de San Diego (USA) et envoyés aux présents auteurs. Le corpus total comprend 24 empreintes de loup sauvage (13 relevés provenant de Pologne, 11 in Harris & Beam 1983), 9 empreintes de cuon captif (6 moulages, 3 photographies) auxquelles ont été intégrées 5 empreintes de canidés de la grotte Chauvet Pont d’Arc ainsi que 8 empreintes de loup du site pléistocène fluviatile de Bottrop, dont les dimensions globales (L x l) ont été reprises20. Pour 3 d’entre elles, les autres mesures ont été calculées à partir des illustrations.
![Correspondance des codes de mesure pour les empreintes de loup. [1] : Harris & Beam (1983), Halpenny (1995) ; [2] :Ledoux & Boudadi-Maligne (2015). L = longueur ; l = largeur.](https://una-editions.fr/wp-content/uploads/2020/09/DAN@_3_Fosse_etal1-300-1024x511.jpg)
[1] : Harris & Beam (1983), Halpenny (1995) ;
[2] : Ledoux & Boudadi-Maligne (2015).
L = longueur ; l = largeur.
Les empreintes de la grotte Chauvet Pont d’Arc ont été étudiées par photogrammétrie et proviennent de l’éboulis d’entrée (secteur EB 18.03), de la galerie du Cierge, de la Salle Hillaire, en amont du panneau du Cheval gravé et de la Salle du Crâne, au pied du panneau des Mammouths raclés. Pour rappel, la photogrammétrie vise à prendre des mesures dans une scène (une photographie ou une scène tridimensionnelle). Pour ce faire, un enregistrement de la scène est réalisé par le biais de photographies de la scène et restituée en trois dimensions grâce à la Corrélation Épipolaire Dense (technique par laquelle il est possible, à partir de plusieurs points de vue d’un même objet, de déduire la position dans un espace relatif de point caractéristique), qui produit un enregistrement fidèle de la réalité. A la grotte Chauvet, les évolutions du protocole d’acquisition sont surtout liées aux nouveaux éclairages utilisés (lampes ballon Airstar Sirocco Led 120W) beaucoup plus puissants ainsi qu’à l’utilisation de boîtiers photographiques de type Ricoh GR II. Les photographies acquises sont par la suite traitées via une suite logicielle (Photoscan21 pour la réalisation des nuages de points denses et des modèles 3D et CloudCompare22 pour l’acquisition des données métriques). Les différentes variables métriques présentées ont ensuite fait l’objet d’une analyse multivariée (analyse en composantes principales). L’ensemble des traitements et analyses statistiques ont été réalisés avec R version 3.6.3 (2020-02-29)23 et des packages complémentaires suivants : dplyr24, FactoMineR25, ggplot226, tidyr27.
Des données ichnométriques générales ont également été recueillies à des fins de morphologie comparative pour le léopard africain28, le lion africain29, le grand félin des cavernes30, l’hyène brune31 et l’hyène des cavernes (Fosse inédit, à partir de 2 empreintes relevées sur élastomère provenant de la grotte d’Aldène ; archives L. Pales au Musée national de Préhistoire et documents personnels de M.A. Garcia).
Les grands Canidés de la grotte Chauvet Pont d’Arc :
données paléontologiques
Initialement organisée en deux axes de recherche juxtaposés, l’approche paléontologique et ichnologique des sols est désormais effectuée, depuis 2016, de manière globale et complémentaire ; des recherches en paléogénétique sur des coprolithes (loup) complètent ces recherches de terrain (J.M. Elalouf).
Les vestiges paléontologiques sont attribués aux canidés selon deux gradients de confiance : les restes accessibles et mesurables présentant des caractéristiques morphologiques et morphométriques assurément de canidés et les spécimens observables à distance, enrobés d’argile (crâne du Belvédère), enchâssés dans les planchers stalagmitique (crâne de la Salle Brunel, éléments postcrâniens de l’Eboulis d’entrée), partiellement mesurables ou difficiles d’accès voire inaccessibles.
À la grotte Chauvet Pont d’Arc, les restes de canidés sont présents depuis l’entrée paléolithique jusqu’à la galerie terminale et confinée du Belvédère, à quelque 200 mètres de l’entrée naturelle (Fig. 2). Les secteurs proches du porche recèlent la majeure partie des vestiges paléontologiques ; dans les autres salles (Salle Brunel, Cactus, galerie Rouzaud, Panneaux Rouges, Salle Hillaire (sondage), Belvédère), l’identification des canidés ne repose que sur un ou deux os par amas / zone.

Le loup
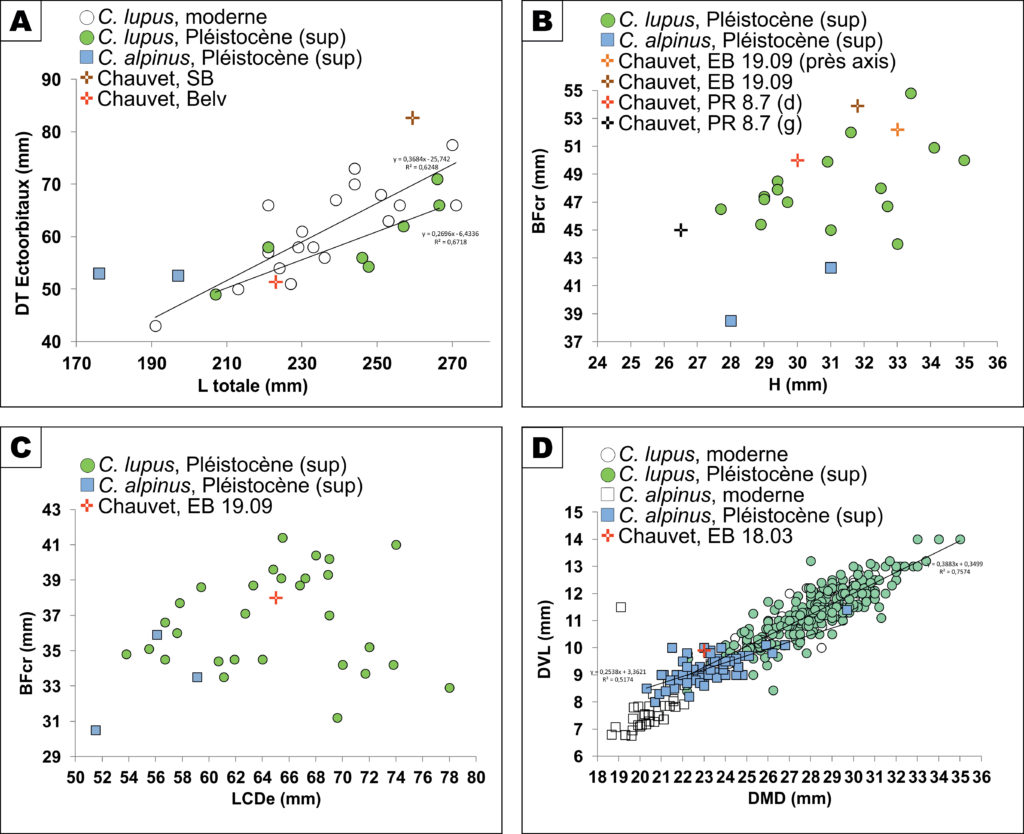
Le crâne scellé dans le plancher stalagmitique de la Salle Brunel, presque complet (absence de l’arcade zygomatique droite), est attribué au loup, d’après les mesures effectuées par photogrammétrie. La longueur totale est de 223,0 mm pour une largeur aux ecto-orbitaux de 51,4 mm. Ces dimensions dépassent systématiquement les valeurs notées sur les spécimens de cuon pléistocène32 et s’insèrent dans la variabilité intraspécifique pour les loups modernes et pléistocènes (Fig. 3a). Le crâne de la Salle Brunel est proche de l’exemplaire de Gerde, rapporté à une louve33.
Voir texte pour explications des mesures.
Le second crâne, niché dans une étroite alcôve terminale de la galerie du Belvédère, présente, en dépit d’un important enrobage d’argile plastique, les caractéristiques morphologiques des canidés (museau effilé, arrière crâne arrondi). Bien que les mesures relevées par corrélation avec une échelle posée au plus près de l’échantillon surévaluent très certainement sa robustesse, ce crâne peut être rapporté à un loup de belle taille, comparable aux plus grands loups modernes et pléistocènes. La longueur totale calculée est de 259,4 mm pour un diamètre transverse au niveau des ecto-orbitaux de 82,7 mm (Fig. 3a).
Sur les 4 atlas identifiés (2 dans la zone de l’Éboulis d’entrée (EB 19.09), 2 juxtaposés dans un amas d’os d’ours des cavernes (PR 8.7) aux Panneaux Rouges, cf. Fig. 2), la hauteur (H) ainsi que le diamètre transverse de la facette crâniale (BFcr) ont été mesurés. Bien que les données ostéométriques soient peu abondantes pour le cuon (n=234), au moins 3 atlas de la grotte Chauvet Pont d’Arc peuvent être rapportés au loup en raison de leurs dimensions plutôt robustes (Fig. 3b). Ces échantillons s’insèrent parmi les spécimens de Malarnaud35 ou de l’aven de l’Arquet (Lateur, inédit). Seul l’atlas PR 8.7(g), présentant une hauteur particulièrement modeste (26,5 mm) se démarque sensiblement du corpus comparatif (n=16) retenu. En l’état (manipulation très restreinte), cette pièce est rapportée à cf. C. lupus.
Pour l’axis, la longueur totale (LCDe) et le diamètre transverse de la facette articulaire crâniale (BFcr) ont été mesurés et comparés aux données recueillies pour le cuon (n=3) et le loup (n=3036) pléistocènes. L’échantillon de Chauvet Pont d’Arc (EB 19.09) est proche des axis recueillis dans des gisements régionaux du Pléistocène supérieur, à l’aven de l’Arquet (Lateur, inédit) ou à l’abri des Pêcheurs (Fourvel, inédit) et s’insère parmi les populations de loup, systématiquement plus robustes que les spécimens attribués au cuon pour le Pléistocène supérieur (Fig. 3c).
Un humérus gauche, réduit à sa portion distale, a été identifié dans ce même secteur de l’éboulis d’entrée (EB 19.09). Le diamètre transverse distal (Bd) est de 39,3 mm et s’insère dans la variabilité intra-spécifique relevée sur de nombreux échantillons attribués au loup, parfois robuste, du Pléistocène supérieur. Enfin, un fémur, pathologique et rongé dans sa partie proximale, est présent dans le secteur EB.04, à proximité immédiate du crâne d’ours n°1. Avec un diamètre transverse distal de 40,8 mm, ce fémur est plus petit que la plupart des spécimens datés des stades isotopiques 3 et 2 (Maldidier, Jaurens37) mais est supérieur au cuon d’Obarreta38. Cette pièce est rapportée à un loup, de taille modeste.
En résumé, le loup semble être l’unique canidé présent dans toutes les galeries de la cavité, à l’exception de 2 secteurs adjacents proches de l’entrée paléolithique : EB 18.03 et EB 27. Des 4 individus adultes décomptés, il pourrait y avoir au moins une louve (Salle Brunel).
Le cuon
Lors de ses recherches en ichnologie, M.A. Garcia avait repéré dans une zone en amont de l’éboulis d’entrée (EB 18.03 – EB 27) de nombreuses empreintes tassées sur une surface d’environ 20 m2 et les avait attribuées à un canidé. Cette plage tamponnée est entaillée de profondes dépressions correspondant à des empreintes d’ours des cavernes ; quelques ossements de canidés (une mandibule gauche complète, deux canines, une carnassière inférieure droite ainsi qu’un calcaneus droit) jonchent le sol. Ce secteur constitue l’un des premiers points ossifères de la cavité mais est difficile d’accès. La reprise des recherches a constitué en une couverture photographique intégrale de cette zone (1300 photos environ en cours de traitement) afin d’obtenir un document de travail photogrammétrique pour l’ichnologie et la paléontologie (repérage et mesures des empreintes et des ossements, chronologie inter- empreintes).
La mandibule gauche, qui repose sur sa face linguale, est recouverte d’un dépôt de calcite cristallin recouvrant toute la face vestibulaire, masquant les foramens mentonniers ainsi que les couronnes de toutes les dents encore enchâssées dans leur support osseux (P/2, P/3, racines de P/4, M/1 et M/2). Les dents sont usées. L’absence de la troisième molaire ainsi que la forme typique du talonide de la carnassière constituent de bons critères de diagnose générique39. Le bord inférieur de la branche horizontale est rectiligne, la base de la branche montante forme un décrochement très net. Le processus coronoide est relativement anguleux et l’incisure mandibulaire est bien marquée. La longueur totale (infradental – processus condylaire) est de 146 mm. Les dimensions restituées de la carnassière (M/1) sont de 23 mm (DMD) x 9,9 mm (DVL) et correspondent à un cuon relativement fort (Fig. 3d), systématiquement plus petit que le loup au Pléistocène supérieur. Des cuons robustes ont été décrits en Italie (Buca del Tasso, Melpignano40), dans des remplissages pro parte synchrones de la grotte Chauvet Pont d’Arc.
D’autres éléments osseux encroûtés ou noyés dans un dépôt de calcite ont été identifiés à 3 mètres environ de ce secteur (EB 27). Une mandibule droite, reposant sur sa face vestibulaire pourrait être appariée à la pièce précédente, en raison de dimensions similaires (Longueur infradental – processus condylaire : 145,3 mm). Cette mandibule scellée au dépôt calcitique est recouverte d’un épais concrétionnement interdisant toute description des dents. Quelques os longs présentent ce même aspect de fossilisation ; la longueur estimée d’une ulna est de 203 mm (185,5 mm sur le spécimen d’Obarreta41 ; systématiquement plus de 250 mm pour le loup42. Le radius, dont les mesures sont approximatives, notamment au niveau de l’extrémité proximale enfouie dans un glacis calcitique, affiche une longueur totale de 168 mm et un diamètre transverse proximal de 24,6 mm. Cet échantillon est à rapprocher des quelques spécimens de cuon du Pléistocène supérieur, qui présentent parfois des dimensions plus fortes (Malarnaud43, El Parpallo44). Enfin une vertèbre caudale et une première phalange, graciles, ont également été rapportées au cuon. Une date radiocarbone est en cours de réalisation sur la phalange (H. Valladas).
En résumé, une petite dizaine d’éléments squelettiques de morphologie et taille similaires provenant de ce secteur proche de l’entrée est attribué au cuon. Ces ossements proviennent probablement d’un seul squelette, disloqué et éparpillé sur quelques mètres carrés et sont déconnectés de l’important bone bed où ont été identifiés l’ours des cavernes, le loup et le bouquetin.
Les grands Canidés de la grotte Chauvet Pont d’Arc :
données ichnologiques
La grotte Chauvet Pont d’Arc renferme de nombreuses empreintes d’ours des cavernes dans toutes les galeries accessibles (éboulis d’entrée, Salle des Bauges, Panneaux Rouges, Salle Hillaire, Salle du Crâne) mais également des empreintes de canidés. A l’exception du secteur EB 18.03, qui se présente sous la forme d’un véritable tapis damé par des dizaines d’empreintes “sédentaires”, toutes les traces sont trouvées sous la forme de spots isolés. À ce jour, au moins 10 zones à empreintes ont été localisées (Fig. 4), correspondant globalement aux observations de M.A. Garcia. Cette répartition couvre l’ensemble des galeries, depuis l’entrée paléolithique jusqu’aux secteurs les plus profonds de la cavité (galerie des Croisillons où ont été identifiées les empreintes humaines45, Sacristie (salle non revue par les présents auteurs). Ces zones ichnologiques se présentent sous forme d’empreintes isolées, lessivées et peu visibles dans les salles à dépôt sédimentaire limoneux (Salle Brunel, Cactus, Panneaux Rouges, galerie des Croisillons) et sous forme de véritables fenêtres ponctuelles sur les paléosols dans les salles où l’argile plastique a conservé ses propriétés physiques et mécaniques et où une calcitation fine a amorcé la fixation des empreintes (galerie du Cierge, Salle Hillaire (panneau du Cheval gravé), Salle du Crâne (panneau des Mammouths râclés)). Ces trois derniers locii constituent les meilleurs spots de relevés ichnologiques ; une couverture photographique a par conséquent été réalisée, dans une perspective photogrammétrique. Ce traitement permet une meilleure individualisation des traces, en raison du recouvrement des empreintes antérieures par les postérieures, de l’effacement de certaines pelotes ou encore de leur orientation différenciée.

Morphologie générale des empreintes
L’identification (spécifique) des empreintes repose sur des critères morphologiques et morphométriques sur une seule trace ou, pour certains carnivores (canidés), préférentiellement sur une piste ou une voie (rectitude, alignements des empreintes).
De manière générale, les félidés (léopard, lion) impriment des traces plus rondes que les autres carnivores (hyénidés, canidés), avec 4 pelotes digitales dépourvues de griffes (rétractiles) et plutôt arrondies ; la pelote palmaire/plantaire forme un M évasé à sa base trilobée alors que son bord antérieur atteint la moitié des pelotes digitales externes (Fig. 5-a1). Les dimensions (L x l) relevées pour les empreintes antérieures et postérieures sont d’environ 6 x 9 cm de côté pour le léopard africain46, 9 x 14 cm pour le lion africain47 et 12 x 15,5 cm pour le lion des cavernes48.

Les empreintes des hyénidés se caractérisent par la présence de 4 pelotes digitales allongées longitudinalement avec griffes marquées et une pelote palmaire/plantaire au contour simple, le bord antérieur comprimé transversalement s’insérant entre les pelotes digitales externes recourbées et le bord postérieur linéaire et concave en son milieu (Fig. 5-a2). Les dimensions maximales des empreintes antérieures et postérieures sont de 9 x 7 cm49 pour l’hyène tachetée (Crocuta crocuta) et l’hyène brune (Parahyaena brunnea) et de 11 x 10 cm pour 2 empreintes mesurées sur élastomère pour l’hyène des cavernes à Aldène.
Les empreintes des canidés (Canis) présentent une morphologie globalement étirée longitudinalement, avec des pelotes non arrondies, de dimensions (L x l) comparables. Chez le loup, les empreintes comprennent 4 pelotes digitales, souvent surmontées par les griffes puissantes des 2 doigts médians50. Les empreintes présentent globalement un aspect “resserré”, les intervalles inter pelotes digitales étant faibles. En revanche, l’écart entre le bord antérieur de la pelote palmaire/plantaire et la partie basale des pelotes digitales médianes (mesure F) est important (Fig. 5-a3). La pelote palmaire/plantaire, parfois comprimée antéro-postérieurement, est étirée transversalement et sa partie basale se compose de 2 “lobes” externes. Le bord antérieur de cette pelote peut arriver jusqu’à la moitié des pelotes externes. Une piste de loup est caractérisée par des empreintes se succédant le long d’un axe rectiligne, les empreintes postérieures mordant sur les antérieures alors qu’une piste d’un chien est formée d’empreintes en zig-zag. Pour le cuon, aucune information sur la morphologie des empreintes n’a pu être trouvée. Cuon et loup ayant une répartition géographique exclusive, les descriptions des empreintes se bornent à la dichotomie canidés – félidés, aucune étude d’ichnologie comparée n’existant apparemment. Quelques remarques laconiques suggèrent des dimensions plus petites pour les empreintes de cuon, conformément aux différences de taille corporelle existant entre les deux canidés. Les empreintes antérieures et postérieures du cuon sont composées de 4 pelotes digitales relativement écartées, allongées (L > l) et rectilignes. Les griffes des 4 doigts sont fréquemment marquées. Les pelotes digitales médianes se démarquent nettement des pelotes externes. Par conséquent, le bord antérieur de ces dernières émarge peu sur la partie basale des pelotes médianes. La pelote palmaire/plantaire est moins ramassée antéro-postérieurement comparativement aux empreintes de loup et la morphologie générale est sensiblement plus arrondie, s’inscrivant dans un ovale nettement moins étiré (Fig. 5-a4). La ligne du bord postérieur semble être moins concave que sur les empreintes de loup.
À la grotte Chauvet Pont d’Arc, les empreintes sont de taille homogène et présentent toutes les caractéristiques décrites ci-dessus pour les canidés. Dans l’éboulis d’entrée, secteur EB 18.03, l’empreinte complète fait face à une main d’ours des cavernes et est dépourvue des griffes médianes. Le voile de calcite blanche amorçant le recouvrement de cet échantillon pourrait expliquer l’absence visuelle des griffes (Fig. 5-b1). La pelote palmaire/plantaire est relativement allongée antéro-postérieurement. Dans la galerie du Cierge, une talus d’argile plastique d’un peu plus de un mètre carré surplombe le sol calcité. Dans cette zone, des empreintes se recoupant partiellement ont été repérées et une empreinte complète et individualisable se démarque nettement (Fig. 5-b2). Les griffes des pelotes médianes sont visibles. Dans la Salle Hillaire, une plage d’argile grise d’environ trois mètres carrés est non recouverte par la calcite a été étudiée par M.A. Garcia (moulages inédits). Ce secteur, situé en amont du panneau du Cheval gravé, comprend au moins 11 empreintes (numéros individuels in situ) pour faciliter le repérage. Ces empreintes correspondent à un mouvement à très faible vitesse vers le fond de la salle, sur un sol irrégulier. Une empreinte, se dégage de cette zone de piétinement et a été isolée pour mesurage (Fig. 5-b3). Dans la Salle du Crâne, plusieurs secteurs ont enregistré les passages des canidés ; à la jonction avec la Salle Hillaire, une empreinte est imprimée dans l’argile comblant une boite crânienne d’ours des cavernes ; dans le prolongement (paroi Sud), sous les pendants aux Rennes menant à des banquettes d’argile, le sol est criblé de dépressions circulaires profondes avec des bourrelets de sédiments épais correspondant à des empreintes d’ours des cavernes (parfois individualisables). Au sommet de ces mottes d’argile, des empreintes de canidés ont été identifiées à plusieurs reprises. La zone où les empreintes sont les mieux préservées se situe à l’aplomb du panneau des Mammouths raclés (paroi Nord). De configuration tout à fait similaire à celle de la Salle Hillaire, une plage d’argile rougeâtre, progressivement reprise par la calcite, laisse émerger une vingtaine d’empreintes parfois très nettes, les griffes des doigts médians et externes ayant imprimé le sol (Fig. 5-b4).
Cette sélection de 5 empreintes de différentes salles, caractéristiques des canidés, sert de support aux observations ichnométriques.
Ichnométrie
Avant d’envisager toute approche comparative entre les registres moderne et fossile, il convient d’explorer les différentes variables métriques retenues (voir Fig. 1). Une première approche strictement confinée aux canidés modernes (loup vs cuon) a été réalisée (Fig. 6). La distribution des différentes mesures laisse entrevoir des distinctions nettes entre les deux taxons, pour certaines variables (mesures E, A, H, C). A partir de ces observations, une comparaison avec les données pléistocènes (grotte Chauvet Pont d’Arc en particulier) a été entreprise, notamment pour 3 variables principales (mesures B, C, D, voir Fig. 1 ; Tab. 2). A première vue, les loups et cuons modernes semblent bien se distinguer par leurs proportions globales (Tab. 3 ; Fig. 7), nettement plus importante chez le premier (environ 10 mm de plus par variable). Seul la largeur maximale des empreintes (mesure D) est indistincte entre loup et cuon (Tab. 2). Les données pléistocènes de Bottrop sont concordantes (et même plus importantes) avec les données des loups modernes. Seule la largeur palmaire accuse des proportions bien plus faibles (comparable au cuon moderne). Il est très probable qu’il s’agisse ici d’une sous-estimation des proportions de la pelote palmaire résultant d’une part d’un effectif faible (n=3) et d’autre part d’une estimation de ces mêmes proportions par les auteurs de la présente étude à partir des figurations (in 51).
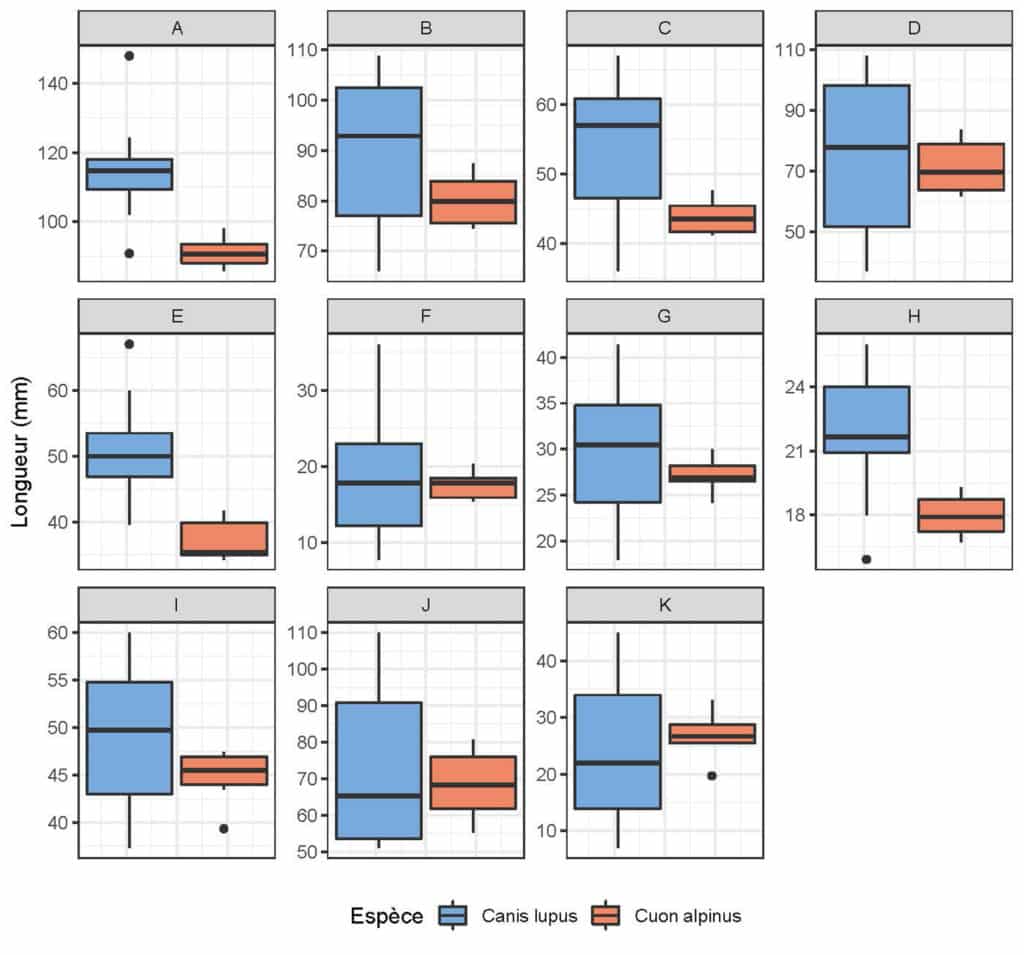
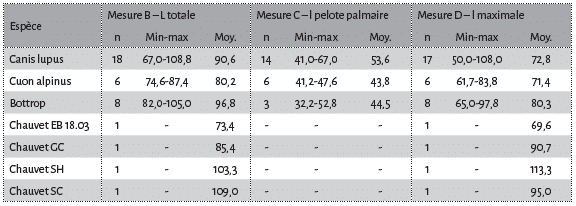
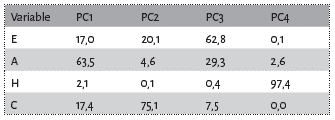
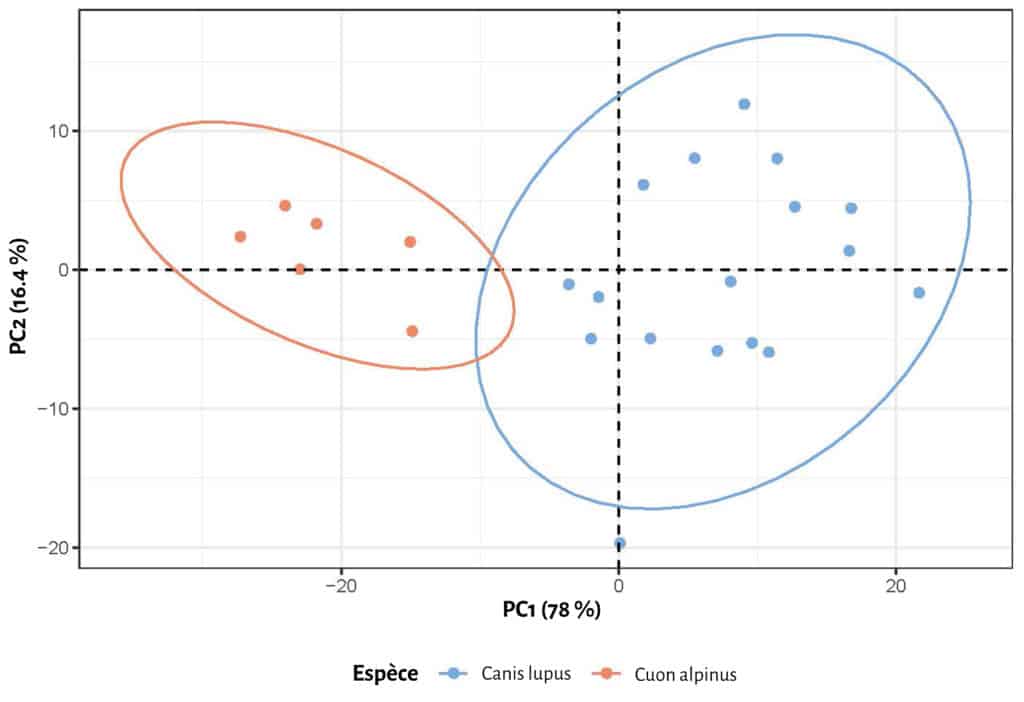
En ce qui concerne les empreintes de la grotte Chauvet Pont d’Arc, 3 d’entre elles s’intègrent très clairement dans le champ de variation d loup (une empreinte de la Salle Hilaire et deux de la Salle du Crâne, Tab. 4 ; Fig. 8). L’ empreinte de l’Éboulis (EB 18.03) accuse de très faibles proportions la rapprochant du cuon. L’attribution à celui-ci semble d’autant plus probable que les plages d’empreintes de ce secteur sont directement associées aux vestiges paléontologiques de la même espèce. L’empreinte de la Salle du Cierge occupe une position intermédiaire, sans savoir, à ce stade, si cet échantillon représente une petite louve ou un cuon de fortes proportions.
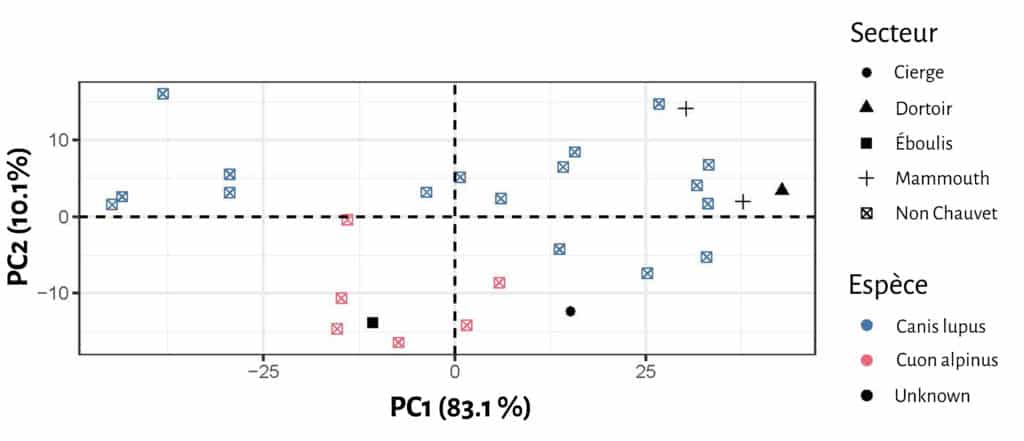

Ces résultats préliminaires, en plus d’autoriser une distinction certaine entre loup et cuon sur la base de variables métriques, permet d’aborder l’identification spécifique d’après l’enregistrement ichnologique. Les premières données issues des empreintes permet de proposer, à l’instar du registre paléontologique que le loup semble avoir fréquenté les secteurs profonds de la cavité, le cuon étant confiné à proximité du secteur d’entrée.
Discussion
La présence conjointe de vestiges paléontologiques et ichnologiques de canidés est rare dans le registre pléistocène.
À l’exception des sites ayant livré des traces de découpe sur les os (Cova Negra, Santa Maira33) et les gisements du Tardiglaciaire a priori exclusivement anthropiques (El Rascaño52, La Riera53, La Vache54), le cuon est pressenti être un occupant naturel des cavités. Sa présence se cantonne dans des secteurs proches des entrées, entre plus d’une dizaine de mètres de profondeur à Bolinkoba55, jusqu’à une trentaine de mètres environ à Amalda56 et à Aitzbitarte III57. À la grotte Chauvet Pont d’Arc, la présence du cuon dans une zone confinée entre 30 et 40 mètres du porche paléolithique confirme la fréquentation naturelle des cavités par cette espèce dans des espaces souterrains encore éclairés naturellement.
Pour le Pléistocène supérieur, le cuon est identifié régionalement dans des niveaux moustériens à Payre, la grotte des Pêcheurs, et dans le Sud-Est à La Baume des Peyrards, dans des sites vraisemblablement synchrones des MIS 5 et 4 à la grotte de Mars (Alpes Maritimes) et la grotte de l’Observatoire (Monaco). A l’Aurignacien, quelques mentions principalement du Sud-Ouest (grotte du Piage dans le Lot, Isturitz (IV) dans les Pyrénées-Atlantiques) ou du Nord de l’Espagne (El Castillo) attestent de la présence de cette espèce, plutôt rare désormais. Au Gravettien et au Solutréen, le cuon ne semble identifié qu’en Espagne (Aitzbitarte III, VIa ; Amalda V, VI ; Bolinkoba, VI ; Lezetxiki II / Aitzbitarte III, II-III ; Bolinkoba, IV). Au Magdalénien, l’espèce n’apparaît qu’exceptionnellement dans les cortèges fauniques (grotte des Orciers en Haute Loire et grotte de La Vache en Ariège, niveau 18 de La Riera dans les Cantabres). Sa présence à la grotte Chauvet Pont d’Arc s’inscrit dans une dynamique biogéographique principalement documentée pour les régions sud-européennes (France, Espagne, Italie58). L’espèce ne peut être retenue en tant que composante importante des guildes de la toundra-steppe.
Les données ichnologiques sont quasi inexistantes pour ce carnivore, aussi bien pour le registre moderne que pléistocène (identification dans les grottes). Le traitement photogrammétrique de la zone de l’Éboulis à la grotte Chauvet Pont d’Arc devrait permettre de proposer rapidement des informations sur les caractéristiques (morphologie, mesures) des empreintes de ce canidé. Dans le cas présent, l’ichnologie est un élément clé de la signature taphonomique sur les occupations des cavités par les carnivores. Cette discipline a un rôle entier dans les reconstitutions sur la fonction des sites pléistocènes. Des échantillons conséquents sont en cours de constitution afin d’intégrer de nouvelles variables à cette première approche ichnométrique comparée. La morphologie étant assez clairement distincte entre empreintes des pattes antérieures (de forme globalement circulaire avec une largeur importante entre les doigts latéraux) et des postérieures (nettement comprimées transversalement et accusant une morphologie ovalaire), une approche morphologique détaillée est à réaliser (morphométrie géométrique). Il en est de même pour la prise en compte du sexage de l’animal, les canidés accusant un dimorphisme marqué dont l’influence sur la taille des empreintes peut a priori être évident.
Les fréquentations naturelles des grottes par le loup sont déduites de découvertes de squelettes entiers, des tanières ainsi que des grottes à ours des cavernes. À Mora Cavorso, un squelette d’une louve morte naturellement dans la cavité a été retrouvé à environ 10 mètres de l’entrée59. Les rares sites identifiés comme tanières témoignent également d’une occupation à proximité des porches (à environ 10 mètres également de l’entrée à Maldidier60 et à Praleitz I (vestibule)61. Des incursions dans les grottes à ours des cavernes ont été relevées et témoignent d’un comportement vraisemblablement nécrophage, parfois très loin des entrées naturelles : à la Caverna Generosa, quelques restes de loup sont présents dans la Saletta, à environ 25 mètres de l’entrée naturelle mais sont absents dans les secteurs plus profonds (“Sala Terminale”)62 ; à Pestera Cioclovina Uscata, des restes de loup pourraient également provenir d’une distance comparable63. Dans les grottes les plus profondes (La Balme à Collomb64, Ursilor65), des restes de loup ont été identifiés à presque 200 mètres des entrées pléistocènes. Des coprolithes de loup ont d’ailleurs été identifiés dans plusieurs grottes à ours des cavernes (Sophiehöhle66, grotte Chauvet Pont d’Arc (J.M. Elalouf)). La présence de restes de loup à la grotte Chauvet Pont d’Arc, même à plus de 150 mètres de l’entrée, pourrait correspondre à des incursions naturelles par ce carnivore. Une localisation systématique des vestiges de cette espèce dans les grottes à ours des cavernes, dans une perspective topographique (distance depuis les entrées pléistocènes, circulations à l’intérieur des cavités), serait néanmoins à rechercher pour confirmer pleinement cette hypothèse.
Au Paléolithique supérieur ancien, les loups sont de grande taille67. Les mesures relevées sur le premier jeu de données ichnologiques à la grotte Chauvet Pont d’Arc ne mettent pas en évidence des dimensions supérieures pour les loups à celles relevées sur des individus modernes sauvages ou captifs. L’identification individuelle des empreintes demeure difficile en raison de la nature du sédiment (degré de dessication de l’argile) ou des mouvements de l’animal (superposition des empreintes). À ce jour, les empreintes présentent une orientation préférentielle depuis l’entrée de la cavité vers le fond et sont situées dans des zones peu propices à des déplacements naturels (plafonds bas, pieds de paroi). En dépit de lacunes spatiales évidentes (inaccessibilité de certains secteurs, empreintes ayant peu marqué les sols), la répartition des empreintes de loup se différencie des aires de circulation des ours(e)s des cavernes, au moins dans les secteurs profonds (accès aux bauges). Le repérage d’empreintes partielles (pelotes digitales, griffes isolées) pouvant être attribuées au loup (critères d’identification ?) a pour objectif de compléter la cartographie des déplacements du loup dans la cavité. Les données sur les empreintes des canidés (morphométrie spatialisation) de la grotte Basura68 devraient être particulièrement intéressantes à étudier dans une perspective comparative.
Conclusion
La grotte Chauvet Pont d’Arc renferme une association unique d’enregistrements paléontologiques et ichnologiques relatifs aux canidés. Ce premier travail vise à jeter les bases d’une recherche méthodologique sur ces deux thèmes, qui présentent un intérêt anthropozoologique certain pour la compréhension de cette cavité ornée d’une part et, au delà, pour le début du Paléolithique supérieur (Aurignacien) d’autre part. La confrontation des données modernes et fossiles (ostéométrie, corpus ichnologique, toporgraphie) permet d’envisager une zonation des occupations par le cuon et le loup. Les critères de reconnaissance et d’identification des empreintes des canidés dans les différentes salles de la cavité (ornées vs non ornées) sert d’étalonnage à l’étude en cours des sols de la galerie des Croisillons, qui a livré une douzaine d’empreintes humaines près desquelles ont été repérées des empreintes de canidés. L’association Homme – Canidés (loup) est progressivement documentée par le registre paléontologique pour le Paléolithique supérieur ; les données ichnologiques, bien que limitées à de très rares gisements, pourraient apporter des informations importantes aux débats actuels sur cette histoire anthropozoologique. La grotte Chauvet Pont d’Arc apparaît comme un gisement important à cette réflexion (sensu M.A. Garcia), mais à partir de nouvelles méthodes d’étude (photogrammétrie), d’une approche multidisciplinaire (combinaison des résultats en archéologie, art pariétal, datations radiocarbone, paléontologie) et de la mise en place progressive d’un référentiel ichnologique (loup, cuon).
Remerciements
Les auteurs remercient Myriam Boudadi-Maligne et Jean Baptiste Mallye pour l’organisation de ce colloque. Des remerciements sont adressés aux personnes et/ou institutions ayant apporté des informations/contributions en ichnologie animale : Jean Jacques Cleyet-Merle (Musée national de Préhistoire) pour l’autorisation déjà ancienne de travailler sur les relevés inédits de L. Pales, Randy Rieches, Allison Alberts, Jane Kennedy, Automn Nelson et Justine Zafran (zoo de San Diego) pour la réalisation de moulages et photos d’empreintes de cuon captif, François Baleux (Traces, Toulouse), Thomas Sagory (Musée d’Archéologie nationale) et l’équipe Chauvet pour l’enregistrement et les premiers traitements photogrammétriques des sols de la grotte Chauvet Pont d’Arc. Les auteurs remercient les rapporteurs qui ont permis d’apporter des précisions importantes relatives à la structuration et la compréhension du présent article. Cette première contribution concernant l’ichnologie animale de la grotte Chauvet Pont d’Arc doit beaucoup aux discussions amicales avec Michel Alain Garcia ; que sa mémoire soit ici honorée.
Bibliographie •••
- AgiSoft PhotoScan Professional (Version 1.2.6) (Software). (2016*). http://www.agisoft.com/downloads/installer/
- Agnolin, F.L., Chimento, N.R., Campo, D.H., Magnussen, M., Boh, D. et De Cianni, F. (2018). “Large carnivore footprints from the Late Pleistocene of Argentina”, Ichnos, 26(2), 119-126.
- Altuna, J. (1981) : “Restos oseos del yacimiento prehistorico del Rascaño”, in : Gonzalez Echegaray, J. & Barandiaran Maestu, I., dir., El Paleolitico Superior de la Cueva del Rascano (Santander), Centro de Investigacion y Museo de Altamira. Monografias, 3,223-269.
- Altuna, J. (1983). “Hallazgo de un cuon (Cuon alpinus Pallas) en Obarreta, Gorbea (Vizcaya)”, Kobie, 13, 141-158.
- Altuna, J. (1986) : “The mammalian faunas from the prehistoric site of la Riera”, in : Straus, L.G. & Clark, G., dir., La Riera Cave, Anthropological Research, 237-274.
- Altuna, J. (1990) : “Casa y alimentacion procedente de macromamiferos durante el Paleolitico de Amalda”, in : Altuna, J., Baldeón, A. & Mariezkurrena, K., dir., La Cueva de Amalda (Zestoa, Pais vasco) ; ocupaciones paleoliticas y postpaleoliticas, colleccion Barandarian, 4,149-192.
- Altuna, J. et Mariezkurrena, K. (2016) : “Estudio de los macromamíferos de la cueva de Aitzbitarte III (zona profunda)”, EKOB, 6, 249-315.
- Ambert, P., Colomer, A. et Galant, P. (2000): “Datations mésolithiques des empreintes humaines de l’étage Cathala de la grotte d’Aldène (Cesseras, Hérault)”, Comptes Rendus de l’Académie des Sciences – Series IIA – Earth and Planetary Science, 331(1), 67-74.
- Argant, A. (2010) : “Carnivores (Canidae, Felidae et Ursidae) de Romain-la-Roche (Doubs, France)”, Revue de Paléobiologie, 29(2), 495-601.
- Aujoulat, N., Geneste, J.M., Archambeau, C., Delluc, M., Duday, H. et Gambier, D. (2001) : “La grotte de Cussac (Dordogne). Observations préliminaires”, Paleo, 13, 9-18.
- Ballésio, R. (1979) : “Le gisement pléistocène supérieur de la grotte de Jaurens à Nespouls, Corrèze, France : les carnivores (Mammalia, Carnivora). 1. Canidae et Hyaenidae”, Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon, 17, 25-55.
- Ballésio, R. et Philippe, M. (1995) : “Les canidés pléistocènes de La Balme à Collomb (Commune d’Entremont-le-Vieux, Savoie)”, Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon, 33, 47-68.
- Baryshnikov, G. (2012) : “Pleistocene Canidae (Mammalia, Carnivora) from the Paleolithic Kudaro caves in the Caucasus”, Russian J. Theriol., 11(2), 77-120.
- Bégouën, C. et Vallois, H. (1928) : Les empreintes préhistoriques, Institut International d’Anthropologie, IIIe session, Amsterdam, 20-29/09/1927.
- Bona, F. (2005) : I depositi del Pleistocene Superiore della Caverna Generosa (Lo Co 2694). Analisi Paleontologica ed interpretazioni Paleoambientali, phD, Universita degli Studi di Milano.
- Bonifay, M.F. (1971) : “Carnivores quaternaires du Sud-Est de la France.” Museum national d’Histoire naturelle (Paris), série C, Mémoires, 21(2), 43-377.
- Boudadi-Maligne, M. (2010) : Les Canis pléistocènes du Sud de la France : approche biosystématique, évolutive et biochronologique, thèse de doctorat Université Bordeaux 1.
- Boudadi-Maligne, M. (2012) : “Une nouvelle sous-espèce de loup (Canis lupus maximus nov. subsp.) dans le Pléistocène supérieur d’Europe occidentale”, C.R. Palevol, 11, 475-484.
- Boudadi-Maligne, M., Mallye, J.B., Langlais, M. et Barshay-Szmidt, C. (2012). “Des restes de chiens magdaléniens à l’abri du Morin (Gironde, France). Implications socio-économiques d’une innovation zootechnique”, Paléo, 23, 39-54.
- Brugal, J.P., Fourvel, J.B. et Fosse P. (2017) : “Premières observations sur les guildes de carnivores de la grotte de l’Observatoire (Principauté de Monaco)”, Bulletin du Musée d’Anthropologie Préhistorique de Monaco, 57, 35-48.
- Castaños, P. et Castaños, J. (2017) : “Estudio de la fauna de macromamíferos del yacimiento de Praileaitz I (Deba, Gipuzkao), in : Peñalver Iribarren et al., éd, La Cueva de Praileaitz I (Deba, Gipuzkao, Euskal Herria) : intervención arqueológica 2000-2009, Munibe, 221-265.
- Castel, J.C. (1991) : Essai d’étude taphonomique de vestiges osseux paléolithiques ; l’exemple de la grotte Maldidier (Dordogne), DEAUniversité de Bordeaux I.
- Clot, A. (1980) : La grotte de la Carrière (Gerde, Hautes-Pyrénées). Stratigraphie et Paléontologie des Carnivores, thèse 3e cycle Université Paul Sabatier Toulouse.
- CloudCompare (version 2.10.1) [GPL software]. (2019). http://www.cloudcompare.org/
- Cordy, J.M. (1983) : “Découverte de Cuon alpinus europaeus Bourguinat dans le Quaternaire de Belgique”, in : Poplin, F., dir., La faune et l’Homme préhistorique : dix études en hommage à Jean Bouchud réunies par François Poplin, Société préhistorique française, Mémoire 16, 49-54.
- Diedrich, C. (2013) : “Extinctions of Late Ice Age cave bears as a result of climate/habitat change and large carnivore lion/hyena/wolf predation stress in Europe”, ISRN Zoology, 2013, doi.org/10.1155/2013/138319, 25p.
- Driesch, v. d., A. (1976) : “A guide to the measurement of animal bones from archaeological sites”, Peabody Museum Bulletins, 1, 1-137.
- Duday, H. et Garcia M.A. (1983). “Les empreintes de l’homme préhistorique. La grotte du Pech-Merle à Cabrerets (Lot), une relecture significative des traces de pieds humains”, Bulletin de la Société préhistorique française, 80(7), 208-215.
- Dufour, R. (1989) : Les Carnivores pléistocènes de la caverne de Malarnaud (Ariège), DESS Université de Bordeaux I.
- Fosse, P. et Philippe, M. (2005) : “La faune de la grotte Chauvet : paléobiologie et anthropozoologie”, Bulletin de la Société préhistorique Française, 102(1), 89-102.
- Garcia, M.A. (2005) : “Ichnologie générale de la grotte Chauvet”, Bulletin de la Société préhistorique française, 102(1), 103-108.
- Gatta, M., Rolfo, M., Petronio, C., Salari, L. et Silvestri, L. (2016) : “Late Pleistocene skeleton of Canis lupus L., 1758 from Grotta Mora Cavorso (Jenne, Latium, central Italy)”, Palevol, 15, 941-949.
- Germonpré, M., Lázničková-Galetová, M. et Sablin, M.V. (2012) : “Palaeolithic dogs and the early domestication of the wolf”, Journal of Archaeological Science, 39, 184-202.
- Ghezzo, E. et Rook, L. (2014) : “Cuon alpinus (Pallas, 1811) (Mammalia, Carnivora) from Equi (Late Pleistocene, Massa-Carrara, Italy): anatomical analysis and palaeoethological contextualisation”, Rend. Fis. Acc. Lincei, 25, 491-504.
- Halfpenny, J.C. (1995) : Wolves in Yellowstone : records since 1967, in : Carbyn, L., dir., Greater Yellowstone predators : ecology and conservation of a changing landscape, 111-116.
- Harlé, E. (1891) : “Note sur des mandibules d’un canidé du genre Cuon”, L’Anthropologie, II, 129-140.
- Harris, R.B. et Ream, R.R. (1983) : “A method to aid in discrimination of tracks from wolves and dogs”, in : Carbyn, L.N., dir., Wolves in Canada and Alaska, Canadian Wildlife Service Report Series, 45, 120-124.
- Iriarte-Chiapusso, M.J. et Arrizabalaga, A. (2015) : “El yacimiento arqueológico de Bolinkoba (Abadiño, Bizkaia). crónica de las investigaciones en la cavidad. secuencia estratigráfica y cronología numérica”, Kobie BAI, 6, 5-88.
- Koenigswald von, W., Sander, P.M. Sander, P.M. et Walders, M. (1995) : “Jungpleistozäne Tierfährten aus der Emscher-Niederterrasse von Bottrop-Welheim”, Münchner Geowiss. Abh. (A), 27, 5-50.
- Le, S., Josse, J. et Husson, F. (2008) : “FactoMineR: An R Package for Multivariate Analysis”, Journal of Statistical Software, 25(1), 1-18. doi.org/10.18637/jss.v025.i01
- Ledoux, L. et Boudadi-Maligne, M. (2015) : “The contribution of geometric morphometric analysis to prehistoric ichnology: the example of large canid tracks and their implication for the debate concerning wolf domestication”, Journal of Archaeological Science, 61, 25-35.
- Ledoux, L. (2019) : “L’ichnologie préhistorique et les traces d’activités au sein des cavités ornées : les grottes de Fontanet (Ariège) et de Cussac (Dordogne)”, thèse de doctorat Université Bordeaux 1.
- Mallye, J.B., Costamagno, S., Boudadi-Maligne, M., Prucca, A., Laroulandie, V., Thiébaut, C. et Mourre, V. (2012) : “Dhole (Cuon alpinus) as a bone accumulator and new taphonomic agent? The case of Noisetier cave (French Pyrenees)”, Journal of Taphonomy, 10(3-4), 317-347.
- Morales, J., Senut, B. et Pickford, M. (2011) : “Crocuta dietrichi from Meob, Namibia: implications for the age of the Tsondab Sandstone in the coastal part of the Namib Desert”, Estudios Geológicos, 67(2), 207-215.
- Mountain, E.D. (1966) : “Footprints in calcareous sandstone at Nahoon Point”, S. Afr. J. Sci., 62, 103-111.
- Okarma, H. (1998) : Le loup en Europe, Orléans.
- Pailhaugue, N. (2003) : “La Faune de la salle Monique”, in : Clottes J. & Delporte, H., dir., La grotte de La Vache (Ariège). I, Les occupations du Magdalénien, CTHS et Réunion des musées nationaux éditions, 73-139.
- Pales, L., Tassin de Saint Péreuse, M. et Garcia, M.A. (1976) : “Les empreintes de pieds humains dans les cavernes. Les empreintes du réseau Nord de la caverne de Niaux (Ariège)”, Archives de l’Institut de Paléontologie Humaine, Mémoire, 36.
- Paulus, M. et Tron C. (1947) : “Étude sur la faune quaternaire de la vallée inférieure du Gard ou Gardon. IV. Les loups”, Bulletin du Museum d’Histoire naturelle de Marseille, VII(1), 1-17.
- Péquart, M. et Péquart S.J. (1960) : “Grotte du Mas d’Azil (Ariège). Une nouvelle galerie magdalénienne”, Annales de Paléontologie, 46.
- Pérez Ripoll, M., Morales Pérez, J.V. Sanchis Serra, A., Aura Tortosa, J.E. et Sarrión Montañana, I. (2010) : “Presence of the genus Cuon in upper Pleistocene and initial Holocene sites of the Iberian Peninsula: new remains identified in archaeological contexts of the Mediterranean region”, Journal of archaeological Science, 37(3), 437-450.
- Petrucci, M., Romiti, S. et Sardella, R. (2012) : “The Middle-Late Pleistocene Cuon Hodgson, 1838 (Carnivora, Canidae) from Italy”, Boll Soc Paleont Ital, 51, 137-148.
- Philippe, M. et Fosse P. (2003) : “La faune de la grotte Chauvet (Vallon-Pont-d’Arc, Ardèche). Présentation paléontologique et taphonomique”, Paleo, 15, 123-140.
- Pirie, T.J., Thomas, R.L. et Fellowes, M.D.E. (2016) : “Limitations to recording larger mammalian predators in savannah using camera traps and spoor”, Wildlife Biology, 22(1), 13-21.
- R Core Team (2020) : R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
- Robu, M. (2015) : The Palaeontology of the MIS 3 cave bear bone assemblage from the Urşilor Cave of Chişcău ; osteometry, palaeoichnology, taphonomy, and stable isotopes. phD, Bucarest, “Emil Racoviţă” Institute of Speleology.
- Robu, M., Mirea, I.C., Petculescu, A. et Constantin, S. (2018) : “Palaeoichnology of an MIS 3 cave bear settlement at Ursilor cave (Western Carpathians, Romania)”, Palaeogeography, Palaeoclimatology, Palaeoecology, 493, 126-135.
- Romano, M., Citton, P., Salvador, I., Arobba, D., Rellini, I., Firpo, M., Negrino, F., Zunino, M., Starnini, E. et Avanzini, M. (2019) : “A multidisciplinary approach to a unique Palaeolithic human ichnological record from Italy (Basura cave)”, eLife 8, e45204, doi.org/10.7554/eLife.45204.
- Skinner, J. D. et Aarde van, R.J. (1981) : “The distribution and ecology of the brown hyaena Hyaena brunnea and spotted hyaena Crocuta crocuta in the central Namib desert”, Madoqua, 12(4), 231-239.
- Soficaru, A., Petrea, C. Petrea, C., Dobos, A. et Trinkaus, E. (2007) : “The human cranium from the Pestera Cioclovina Uscata, Romania. Context, age, taphonomy, morphology, and paleopathology”, Current Anthropology, 48(4), 611-619.
- Stander, P.E., Ghau, II., Tsisaba, D., Oma, II. et Vi, I.I. (1997) : “Tracking and the interpretation of spoor: a scientifically sound method in ecology”, Journal of Zoology, 242(2), 329-341.
- Suire, C. (1969) : Contribution à l’étude du genre Canis d’après les vestiges recueillis dans quelques gisements pléistocènes du Sud-Ouest de la France, thèse 3e cycle Université de Bordeaux I.
- Vialou, D. (1979) : “Grotte de l’Aldène à Cesseras (Hérault)”, Gallia Préhistoire, 22(1), 1-85.
- Viehmann, I. (1973) : “Les traces de vie de l’ours des cavernes (Ursus spelaeus) dans les grottes de Roumanie”, Institut de Spéléologie “Émile Racovitza” Livre du Cinquantenaire, 451-461.
- Wickham, H. (2016) : ggplot2: Elegant Graphics for Data Analysis, New York.
- Wickham, H., François, F., Henry, L. et Müller, K. (2020) : dplyr: A Grammar of Data Manipulation. P ackage R version 0.8.5. https://CRAN.R-project.org/package=dplyr
- Wickham, H. et Henry, L.(2020) : tidyr: Tidy Messy Data. Package R version 1.0.2. https://CRAN.R-project.org/package=tidyr
Notes
- Garcia 2005.
- Viehmann 1973 ; Robu et al. 2018.
- von Koenigswald et al. 1995 ; Agnolin et al. 2018.
- Mountain 1966 ; Morales et al. 2011.
- Bégouën & Vallois 1928.
- Péquart 1960.
- Pales et al. 1976.
- Duday & Garcia 1983.
- Aujoulat et al. 2001 ; Ledoux 2018.
- Vialou 1979 ; Ambert et al. 2000.
- Romano et al. 2019.
- Philippe & Fosse 2003 ; Fosse & Philippe 2005.
- Boudadi-Maligne et al. 2012 ; Germonpré et al. 2012.
- von den Driesch 1976.
- Paulus & Tron 1947 ; Suire 1969 ; Bonifay 1971 ; Ballésio 1979 ; Clot 1980 ; Dufour 1989 ; Boudadi-Maligne 2010 ; Argant 2010.
- Harlé 1891 ; Ballésio 1979 ; Altuna 1983 ; Cordy 1983 ; Perez-Ripoll et al. 2010 ; Baryshnikov 2012 ; Petrucci et al. 2012 ; Ghezzo & Rook 2014 ; Brugal et al. 2017.
- Harris & Beam 1983.
- Halfpenny 1995.
- Ledoux & Boudadi-Maligne 2010.
- Agnolin et al. 2018
- AgiSoft PhotoScan Professional 2016.
- CloudCompare (v.2.10.1) 2019.
- R Core Team 2020.
- Wickham et al. 2020.
- Le et al. 2008.
- Wickham 2016.
- Wickham & Henry 2020.
- Stander et al. 1997 ; Pirie et al. 2016.
- Stander et al. 1997.
- Agnolin et al. 2018.
- Skinner & van Aarde 1981.
- Altuna 1983 ; Perez-Ripoll et al. 2010.
- Clot, 1980.
- Altuna 1983 ; Baryshnikov 2012.
- Dufour 1989.
- Suire 1969 ; Dufour 1989 ; Argant 2010 ; Altuna 1983 ; Baryshnikov 2012.
- Boudadi-Maligne 2010.
- Altuna 1983.
- Dufour 1989 ; Harlé 1891.
- Petrucci et al. 2012.
- Altuna 1983.
- Boudadi-Maligne 2010.
- Dufour 1989.
- Perez-Ripoll et al. 2010.
- Garcia, 2005.
- Stander et al. 1997 ; Pirie et al. 2016.
- Stander et al. 1997.
- Agnolin et al. 2018.
- Skinner & van Aarde 1981.
- Okarma 1998.
- Agnolin et al. 2018.
- Altuna 1981.
- Altuna 1986.
- Pailhaugue 2003.
- Iriarte-Chiapusso & Arrizabalaga 2015.
- Altuna 1990.
- Altuna & Mariezkurrena 2016.
- Dufour 1989 ; Altuna 1983 ; Perez-Ripoll et al. 2010 ; Petrucci et al. 2012 ; Ghezzo & Rook 2014 ; Brugal et al. 2017 ; Mallye et al. 2012.
- Gatta et al. 2016.
- Castel 1991.
- Castaños & Castaños 2017.
- Bona 2005.
- Soficaru et al. 2007.
- Ballésio & Philippe 1995.
- Robu, 2015.
- Diedrich 2013.
- Suire 1969 ; Boudadi-Maligne 2010.
- Romano et al. 2019.
