




Introduction
Animal domestication induces not only biological changes of the targeted populations but also cultural shifts in the way the domesticated populations are perceived1. The grey wolf (Canis lupus) was certainly the first animal to be involved in this bio-cultural transformation, leading to the appearance of the dog (Canis familiaris)2, although the precise timing of this change is still intensely debated3. Regardless, during the Late Upper Palaeolithic, alterations in the animal’s morphology4 and modifications in the behaviour of humans and “proto-domesticated dogs” alike5 consensually characterise the early steps of dog domestication. Once domesticated, the dog accompanied human groups in their migrations across the world until regions where the wolf has never been present, such as Australia6 and South America7.
However, this strong relationship never became exclusive and human societies maintained strong economic and symbolic interactions with the dog’s wild relatives. In Europe, the red fox (Vulpes vulpes) has been used, by turns, as a companion8 and a fur provider9. Archaeological evidence also points toward the symbolic consideration of the grey wolf in many cultures across the Northern hemisphere, as a participant in ritual paraphernalia10. Nonetheless, all the studies aiming at precisely reporting the distinct role of dogs and non-dog canids in the archaeological record have been hampered by the difficulty in identifying the different canid species from their osteological remains11.
With 18 to 2012 identified species13, the American continent hosts the largest diversity of canid species in the world. The multiplicity of biologically closely related species increases both the potential of interaction with human societies and the ambiguity in their skeletal identification. In Mesoamerica14, four different species are present since the beginning of the Holocene: the grey fox (Urocyon cinereoargenteus), the coyote (Canis latrans), the Mexican wolf (Canis lupus baileiy) and the dog15. Because of its very small size (on average 3-7 kg for a body length of 1 m including the tail16), the grey fox can often be disentangled from the other species, even from fragmented bones. As a matter of fact, this species is often identified in the archaeozoological record, albeit never in large proportions17. It is particularly scarce in the iconography18 and, unlike the other canids, the fox does not stand as a particular symbol in the Mesoamerican cosmogony (see Cultural background). Therefore, we focused our study on the identification of the three other species, namely the dog, the wolf and the coyote.
In this paper, we aim to deepen our understanding of the role of large canids (Canis spp.) in Mesoamerica by providing a more accurate identification of the canid remains to the species level. After introducing the ecological and cultural background of each animal, we first present a geometric morphometric (GMM) analysis of the first lower molar and test its efficiency to discriminate specimens of known species using a reference collection of 42 modern and archaeological specimens. This approach is then applied to 22 archaeological teeth, each from a unique individual, from four archaeological sites in central Mexico. The subsequent identification is used to contrast our perception of canid diversity and their interaction with human societies in the region.
Ecological background
It is now widely accepted that the dog has been introduced to America after its domestication from Eurasian wolves19 while no evidence for the local domestication of a population of American wolves has been identified so far. Genetic evidence indicate that, outside of the Arctic region, the American dogs evolved in a distinctive monophyletic clade20. It is only after 1492, with the European colonisation and the introduction of large numbers of Eurasian dogs, that this lineage faded, until becoming virtually absent in modern populations, including in breeds of American origin21. In pre-Columbian Mesoamerica, carbon and nitrogen stable isotope analysis indicates that dogs often show similar diets to those of humans22. This trend has been interpreted as an evidence of their close relationship, whether dogs would have been fed purposely on household diet or were scavenging food waste and human faecal material23. In contrast, some individuals with more distant diets also suggest the presence of likely feral dogs, or at least loosely managed ones, around the settlements24.
The Mexican wolf is the smallest subspecies of grey wolf25 and genomic analyses have shown a clear distinction from the other North American populations26. Once inhabiting the dry pine-oak forests ranging from the Southern United States to the isthmus of Tehuantepec27, the subspecies was extinct in the wild by 198028. Based on seven (captive) founder individuals, an intensive programme of captive breeding, management and release led to a new wild population of 21 Mexican wolves in Northern Mexico and around 100 in Arizona and New Mexico, according to a 2015-2016 census29. Reports from the beginning of the 20th century indicate that the Mexican wolves were hunted to extinction to protect livestock30. Nevertheless, studies of scats from wild Mexican wolves in the Blue Range Wolf Recovery Area (spanning between Arizona and New Mexico) show that their natural diet is centred on large preys, more specifically elk (Cervus elaphus) and deer (Odocoileus spp.)31.
The coyote is a widespread Nearctic canid. Whereas Pleistocene deposits have yielded evidence of its presence as far south as Costa Rica32, climate change following the Late Pleistocene would have led to the contraction of its natural distribution to the north33. The coyote is particularly well adapted to prairies and human-modified environment, and during the last centuries, land clearance and the extirpation of its major predator, the wolf, have allowed the species to colonise most of North America and to extend across Central America34. In central Mexico, the coyotes feed mostly on small to medium preys (Lagomorpha, Rodentia), but also fruits during the wet season35, which would have all been available in the vicinity of ancient Mesoamerican settlements.
Due to habitat restriction and population depletion, hybridisation between coyotes and dogs36 as well as wolves and coyotes37 appear to be quite frequent amongst modern North American populations. Besides, pre-contact American dogs share some alleles with modern coyotes and North American wolves, indicating that this admixture goes back to prior 149238. Whilst the spatiotemporal extent of this hybridisation is not well understood yet, it may have played a role in the characterisation of wild and domestic populations alike.
Cultural background
Dogs and humans are closely associated in the Mesoamerican cosmogony. A Mexica belief, reported by the Spanish, tells us that only a dog was able to guide the spirit of the deceased through the death realm and to help him cross the Chiconahuapan river39. The presence of dog remains40 and dog effigies41 in human burials, or dog graves associated with funerary structures42 have all been interpreted as an evidence of those beliefs. In the Mexica mythology, Xolotl, the sky demon responsible for the movements of the stars, was represented by a dog43. But the presence of the dog in daily life is also illustrated by the rare architectural models characteristic of the Late Preclassic (300 a.C. – 300 p.C.) in Western Mexico. These complex ceramic objects represent domestic and village scenes where dogs are often depicted44. Testimonies of the early Spanish conquerors relate the consumption of dogs amongst the populations of central Mexico45, which is confirmed by recurrent presence of dog bones in Mesoamerican middens, sometimes presenting typical patterns of burning or cut marks46.
The identification of other canid species is more heterogeneous. Whether looking at the iconography or the ethnohistoric records, there does not seem to be a straight discrimination between coyotes and wolves in Mesoamerica. In his Historia General de las Cosas de Nueva España, Fray Bernardino de Sahagun introduces the coiotl, an animal close to both the wolf and the fox but neither one of them47. He describes it as a fierce and resentful animal that does not hesitate to attack people and their domestic animals. Its Nahuatl name, coiotl, is at the origin of the modern term “coyote”. A similar animal, only depicted in the Nahuatl version of this ethnohistoric work is the cuitlachtli that could correspond to the wolf48. Perhaps because of the transparency of the term coiotl, historians and archaeologists often identify the non-dog canids present in the Mesoamerican iconography and folklore as coyotes rather than wolves, even though no morphological or ethological reasons are given49.
However, the coyote is particularly discrete in the archaeozoological record: our review of the literature found only six occurrences. Two individuals have been identified in the superficial layers of Cueva de las Varillas, in Teotihuacan, and interpreted as natural accumulation50. Still in Teotihuacan, one coyote has been identified in burial 6 of the Moon Pyramid51. In Western Mexico, on the site of Malpaís Prieto, a complete radius found in the midden of an elite house is particularly slender, and it was identified as a possible coyote52. Finally, two potential individuals (cf. Canis latrans) are mentioned in the Late Postclassic and Colonial occupation of Tipu, Guatemala53. Nonetheless, many authors maintained their identification to the level of the genus (Canis sp.), acknowledging the possible presence of coyotes54.
Without being largely more common than the coyote yet, the wolf is more frequent in the archaeozoological record and always associated with extraordinary deposits. In Teotihuacan, in the dedicatory burials of the Moon Pyramid (burial 2, 4 and 6), the remains of at least 22 wolves were present55, and others were found in the Quetzalcoatl Pyramid, although there is no count56. Several skeletons of wolves were also recovered in the ancient city of Mexico Tenochtitlan57, including two complete individuals (offerings 120 and 125) richly adorned with green-stone beads, shell pendants and gold and copper bells58. Other findings include various remains from tronco-conical pits in Cuanalan, Basin of Mexico59, and a single isolated mandible in a ceremonial cache in Cantona, Puebla60.
It appears from this brief overview that the dogs, the coyotes and the wolves present distinct behaviours and ecological characteristics ranging from domestic (dogs) to commensal (coyotes) and wild animals (wolves). This gradient might be reflected in their cultural perception in ancient Mesoamerica, denoting their relative proximity to the anthropized environment and hence human settlements. Therefore, a more systematic identification of the remains in the archaeozoological record would enhance our understanding of the relationships between human societies and canid populations.
Tooth geometric morphometrics: testing a “new” tool for Canid species identification
Geometric morphometrics (GMM) is an approach allowing the precise capture of phenotypic variations61. Because, in mammals, the shape of each tooth is strictly controlled by complex genetic signals62 and this tissue is hardly ever remodelled through the lifetime of an individual63, tooth morphometry is believed to retain a strong taxonomic and phylogenetic signal. Indeed, recent GMM application to archaeological teeth of rodents64, pigs65 or equids66 have shown the potential of this approach to refine taxonomic identification of ancient remains and unravel population history. Yet, while canid complete skulls and mandibles have been the object of in-depth GMM analyses67, single-tooth morphometrics have been studied less intensively68.
Reference collection
In order to distinguish between the three species of Mesoamerican large canids, we developed a two dimensional GMM protocol on the first lower molar. Forty-two teeth (one per individual) of known species were first analysed (Tab. 1). They consist in 16 modern coyotes, 10 modern wolves, 8 modern dogs and 8 archaeological Mesoamerican dogs genetically identified in previous studies69. All specimens were adults with fully erupted first molars.
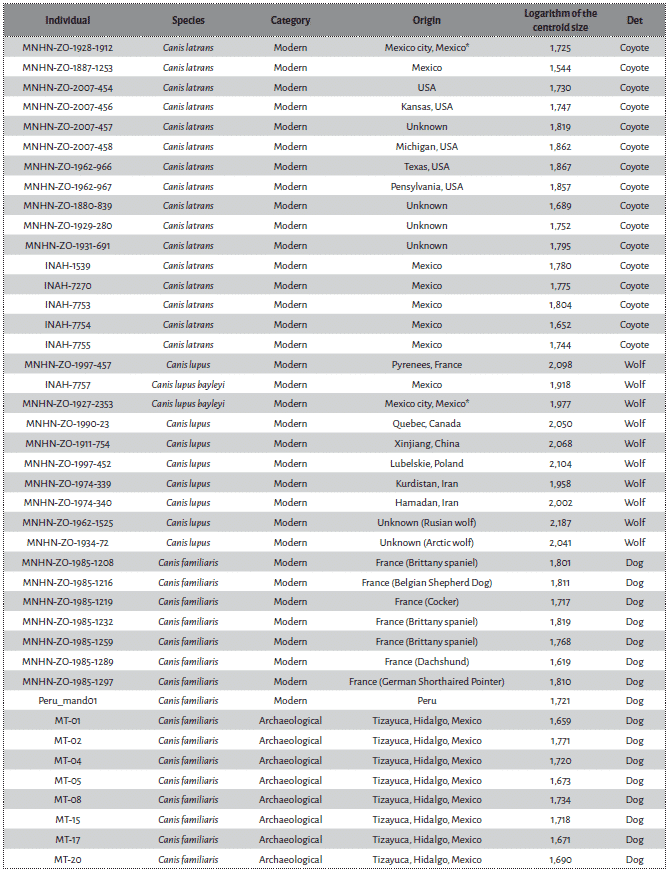
* : Captive individuals from the Zoologico de Chapultepec.
When selecting the wolves, priority was given to the Mexican subspecies, whether they were wild or captive (N=2). A very limited number of individuals was available within this category70, which can be related to the scarcity of the population in the wild. Thus the sampling was extended to wild individuals of known provenience (N=8). Individuals which skull presented “dog” characteristics such as a steep angle between the forehead and the snout71 and a larger orbital angle72, were considered as questionable identification and not used for this study. Similarly, Eastern and red wolves were not considered in this study as they may represent different degrees of hybridisation between wolves, coyotes, and dogs to a lesser extent73.
Because they would match the expected size and robustness of ancient Mesoamerican dogs74, seven modern dogs used as reference were chosen amongst dolicocephalic / mesocephalic and medium-sized breeds. One village dog from Northern Peru, affected with canine ectodermal dysplasia (CED), was also included. Although this congenital defect also impacts the teeth morphology75, all the landmarks used in our protocol were visible on this individual. Because our archaeological collection included dogs potentially affected by CED76, it appeared particularly useful to consider this variation in the tooth morphology. Finally, we used eight archaeological dog mandibles from Tizayuca, Basin of Mexico, that were previously identified as dogs based on their metrics and mitochondrial DNA77.
Archaeological samples
A total of 22 teeth of unidentified canids from four archaeological sites of central Mexico (Fig. 1) was analysed for species identification. All the teeth were fully mineralised and showed limited wear. All the landmarks used in our protocol were visible on each of these teeth.
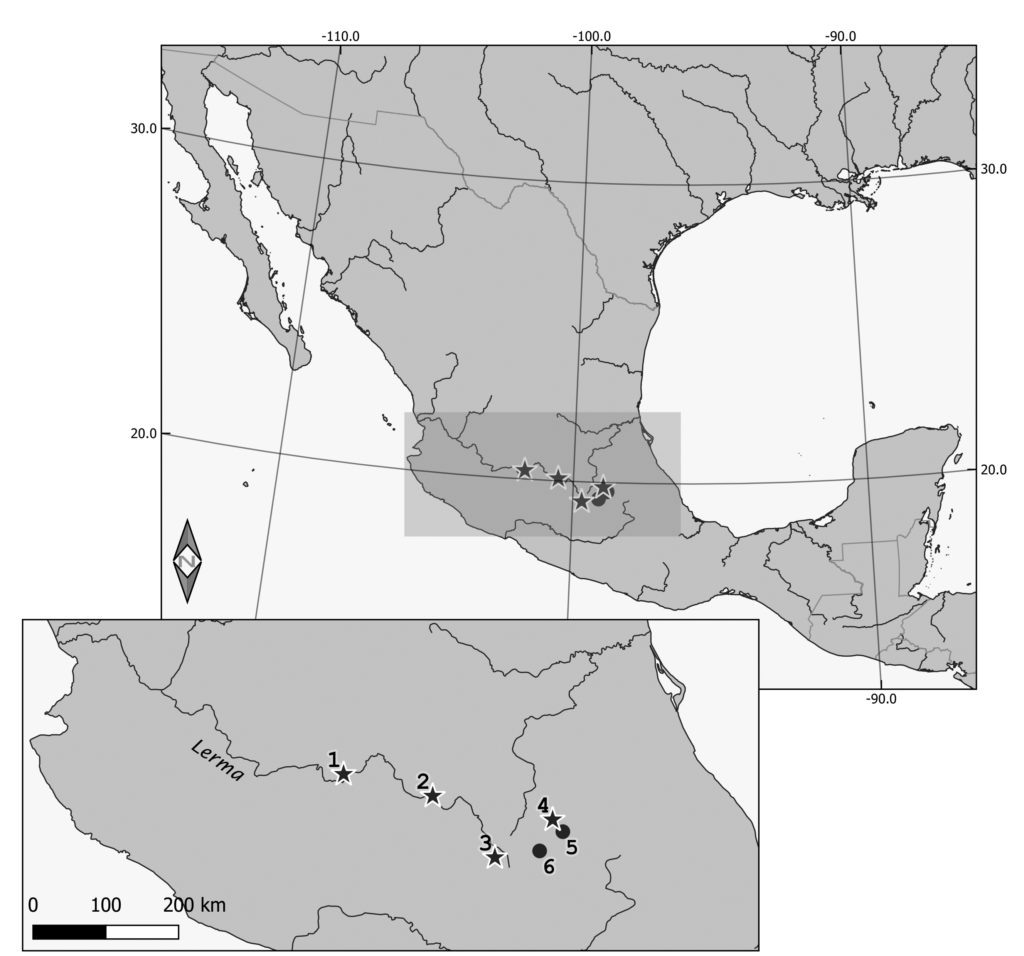
The site of Nogales represents the western end of the collection analysed in this study. It was occupied by sedentary agriculturalists from ca. 500 p.C. until its abandonment around 1000 p.C.; a brief re-occupation by groups of hunters is noted ca. 1450-1500 p.C.78. A fragmented mandible analysed in this study comes from the early occupation of the site, around 500 p.C., and was found in the filling of a platform. The other two remains were isolated teeth found in disturbed layers.
The site of El Mezquite – Los Azules (JR74) is located in the cradle of the Chupicuaro culture, in the Lerma basin79 and was occupied from ca. 600 a.C. to 400 p.C.80. The sample analysed in this study comes from an isolated mandible found in the filling of a Chupicuaro structure dated to the Classic period, between 250 p.C. and 400 p.C.81.
Calixtlahuaca is a large urban centre located in the Toluca valley and occupied between ca. 1100 and 1530 p.C. Historical documents indicate that the city was conquered by the Aztecs of the triple alliance between 1475 and 1478. In 1530, in order to establish their authority on the valley, the Spaniards evacuate the city and relocate its inhabitants in Toluca82. The zooarchaeological analysis shows that the canids were the taxonomic group the most represented in the site, both in terms of NISP (88; 55% of the total NISP) and MNI (5; 24 % of the total MNI)83. The two samples analysed here come from structure 307, the domestic midden of a high-status residence associated with the Ninupi phase (1380-1450 p.C.).
Tizayuca is a settlement located in the basin of Mexico. Architectural, lithic and ceramic remains from the Teotihuacan complex, Toltec and Aztec cultures, as well as sporadic Colonial elements indicate that the area was occupied persistently from 200 to 1520 p.C. However, the stratigraphic layers were particularly thin and prevented the clear chronological attribution of the bone remains84. As in Calixtlahuaca, the canids outnumbered the other taxa (NISP = 877, or 25 % of the total NISP; MNI = 42). Three adults and one juvenile canids were found in burials, the adults being all identified as dogs on the basis of their mitochondrial DNA85. Sixteen first lower molars of unidentified canids were analysed in this study.
Method
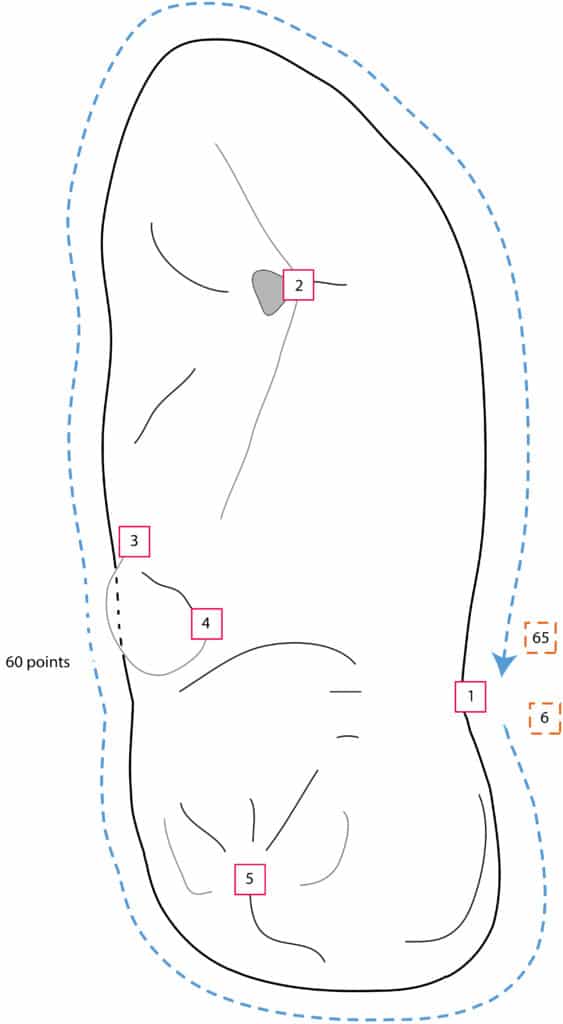
Each tooth was photographed on the occlusal view using a standardised protocol. The teeth were positioned with the talonid being on a horizontal plan and photographed using a Canon EOS 1000D digital camera equipped with a 70-300 mm macro lens. Tooth shape was assessed by digitising five landmark and 60 sliding semi-landmark coordinates on the images (Fig. 2) using tpsDig2 v2.1986. Superimposition and subsequent statistical analyses were performed using R 3.5.387. The coordinates of the different specimens were superimposed and scaled using the Morpho package88. Size analyses were based on the log-transformed centroid size and shape analyses were based on the Procruste residuals (coordinates after superimposition). Differences in size were tested using Kruskal-Wallis and Wilcoxon tests and visualised by boxplots. Analysis of shape variation was based on a principal component analysis (PCA), before testing the differences between groups using multivariate analysis of variance (MANOVA) and linear discriminant analysis (LDA) after reduction of the dimensionality of the data89. The LDA was paired with a leave-one-out cross validation assessing the discriminant power of the analysis. The threshold to confidently identify an individual was set to 95%. Identifications made with a probability of 75 % to 95 % were only considered as plausible.
Statistical tests were performed using a reference threshold of α=0.05 on R v3.5.390.
Identifying the canid species present in Central Mexico
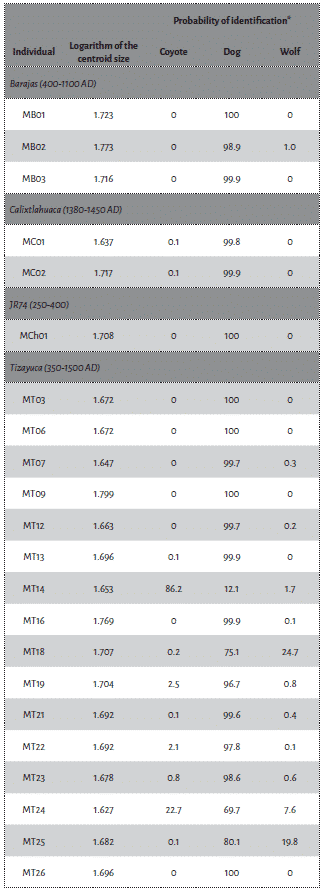
Coyotes, dogs an wolves from the reference dataset differ in size (Kruskal-Wallis test, H=23.5, df=2, p=8e-6), although when they are compared two by two only the wolf differs from the others (Wilcoxon test, p=3.8e-7 for both comparisons), while the coyote does not differ in size from the dog (p=0.17) (Fig. 3). The molar log-transformed centroid size of the wolf ranges from 1.918 to 2.187, that of the coyote from 1.554 to 1.867 and that of the dog from 1.619 to 1.819 (Fig. 3). The molar size of the unknown archaeological samples ranges from 1.637 to 1.799 (Tab. 2) overlapping the sizes of the dogs (p=0.052) while being slightly smaller than the coyote (p=0.0016) and considerably smaller than what we observed in wolves (p=1.9e-7) (Fig. 3).

The first two axes of the shape PCA (21.55 % and 15.96 % of total variance respectively, Fig. 4), tend to separate wolves from the other groups. While coyotes and archaeological unidentified specimens are clearly separated from the wolves, the reference group of dogs (including both modern and archaeological) overlap with wolves and coyotes along axis one, and mostly with coyotes along axis 2. The unidentified archaeological specimens clearly overlap with the coyotes and the dogs, and not a single one overlap with the wolves.
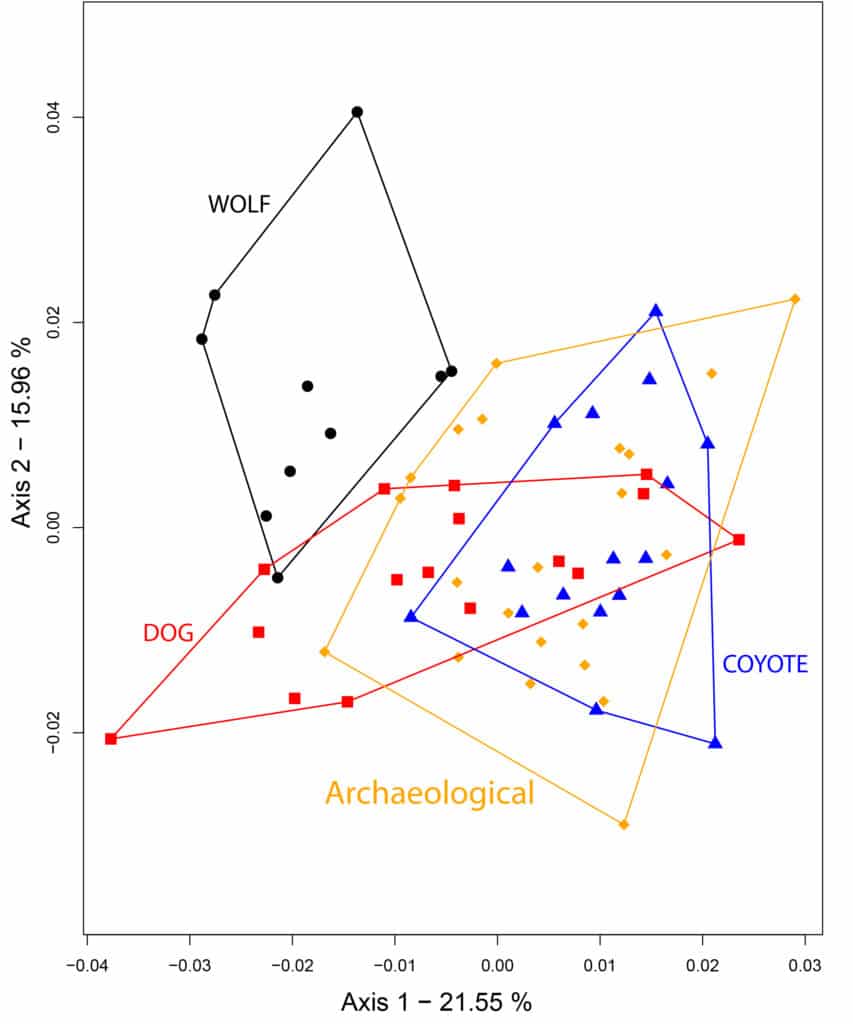
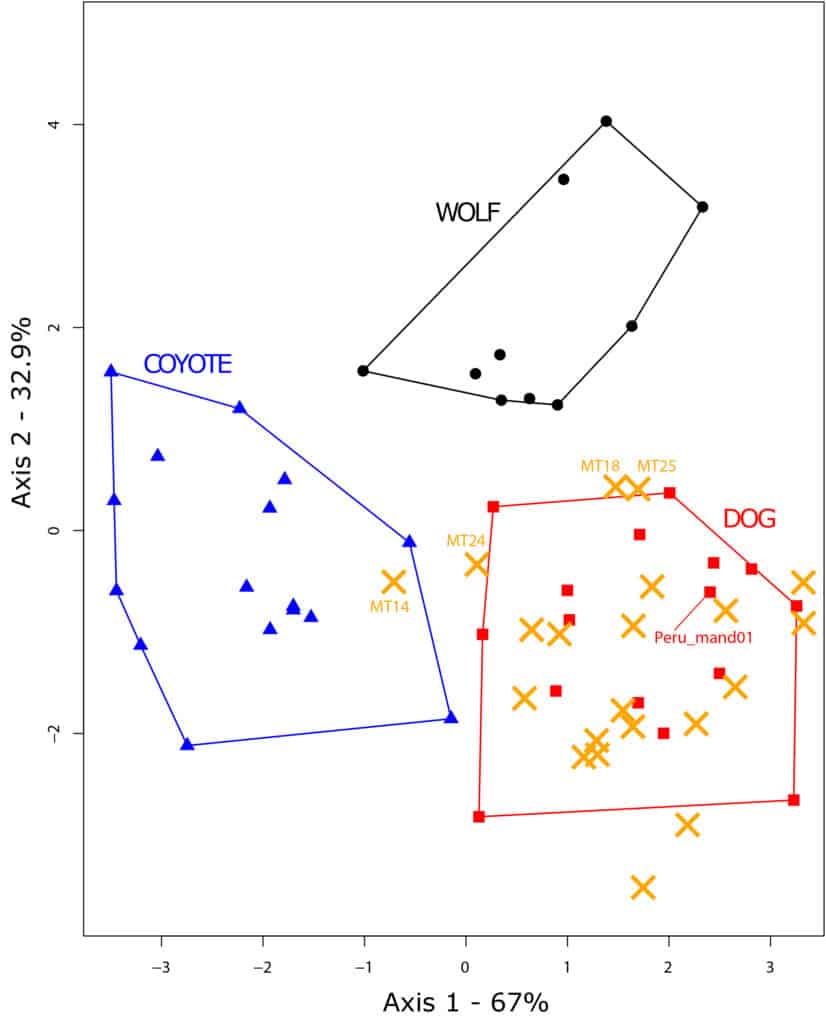
The three reference taxa differ in their first lower molar shape (F(10, 72)=16.369, p=6e-15), and the discriminant analysis reach 88.3 % of correct cross validation. The dog and the coyote are differentiated along the first discriminant axis (67.1 % of the total variation, Fig. 5) while the wolf differentiates along the second component (32.9 % of the total variation). Predicted position of most of the archaeological unknown specimens overlaps with the dog except one (fig. 5). Among the unknown archaeological specimens, 18 dogs were identified with a probability ranging from 96.7 % to 100 % (Tab. 2). Lower probabilities of identification led to the identification of two ‘possible dogs’ with probabilities of identification of 75.1 % (MT18) and 80.1 % (MT25) and one ‘possible coyote’ (MT14) identified with a probability of 86.2 %. Finally, the last specimen (MT24) was identified with a probability of 69.7 % to dog, 22.7 % to coyote and 7.6% to wolf and was left unidentified (Tab. 2).
Discussion
This preliminary study confirms that the GMM analysis of the first lower molar is a promising tool for the taxonomic identification of North American canids. From a set of reference individuals, we show that coyotes, dogs and wolves can be successfully discriminated through their size and shape using a 2D landmark-based approach. Applying this same approach to archaeological samples allows us to discuss more in depth the species distribution and its cultural and ecological implications.
Dogs
Most of the archaeological first lower molars analysed in this study securely come from dogs (18 (+2 possible dogs) out of 22). This result is consistent with previous zooarchaeological studies showing the constant presence of dogs in central Mexico91. In fact, all the individuals from Nogales, JR74 and Calixtlahuaca present a molar morphometry consistent with dogs. The identification in Tizayuca is, however, more problematic. Twelve individuals are confidently identified as dogs whereas two individuals could only be identified as possible dogs due to their lower probability of identification. This variation in the probability of identification may be related to a more diverse tooth shape spectrum in the dogs from Tizayuca. Yet, the dogs from this site show diverse morphotypes, including the presence of possible hairless dogs92. The hypothesis that some individuals might be the result of hybridisations between dogs and wolves or dogs and coyotes is also considered bellow.
Coyotes
Although coyotes are quite rare in the archaeological record of central Mexico, one individual from Tizayuca have been identified as a possible coyote on the basis of its tooth shape, with a probability of 86 %. The presence of a coyote in this specific site could be related to the high number of faunal remains retrieved during the archaeological exploration (more than 3000, including 877 canid remains), as a larger amount of bones would increase the probability of identifying scarce or new species93. The large number of canid individuals identified in Tizayuca and their prevalence on the site also suggest a major interest for that taxonomic group, whether it was for economic or symbolic purposes. However, while dogs have been found in burials, associated with civic-ceremonial or domestic spaces, the possible coyote mandible was found as isolated and fragmented, in a pit, with very limited contextualisation.
Wolves
No wolves were confidently identified in our study, although they have been reported in other sites in the region, in particular Teotihuacan and Mexico-Tenochtitlan94. In these two sites, their bodies were found in exceptional offerings where they took part in state-level rituals95. As none of our samples come from such symbolic context, the absence of wolves is consistent with their peculiar cultural association.
Hybridization between dogs and wild canids?
Evidence of genetic admixture between dogs and wolves or coyotes has been found in the archaeological record96 and amongst modern populations97. Discovering the bones of hybrids of dogs and wolves, or coyotes, in the archaeological record is thus plausible. As a matter of fact, some authors have suggested the identification wolf-dog hybrids based on morphoscopic and metric analyses, assuming that first generation hybrids would inherit a mixture of characters from each parent98. Nonetheless, the relationship between genotype and phenotype is not strictly cumulative, in particular due to epistatic phenomena where a combination of genes and alleles will interact to produce a specific phenotype. Experimentations have shown that if some hybrids tend to present an intermediate morphology99, others display an original phenotype100. Questionable specimens MT18 and MT25, and above all MT24, might be the evidence of a significant admixture between dogs and wild canids. However, addressing this question would require to expand the reference collection in three directions. First of all, the use of a larger reference collection for each species is required to ensure the diversity of tooth shape is adequately captured in the analysis. Secondly, the inclusion of known hybrid populations such as the Eastern and the red wolves, in large enough numbers, would provide an overview of the diversity in such groups. Finally, the degree of admixture in these hybrid individuals should be quantified using genetic tools, in order to identify the range of morphological variations in connection with relative dog, wolf and coyote ancestry.
Ecological and cultural impact of the distribution of canids in the archaeological sites of central Mexico
Through this study, we highlight the broad distribution and the prevalence of the dog amongst the canids present in the zooarchaeological record of central Mexico. By its absence in the corpus analysed here, the highly symbolic role of the wolf is comforted. Although it is hard to infer past distribution of a species from its modern distribution, the wolf’s ecology and behaviour implies that it would not have been found regularly near human settlements but rather in forested habitats. There are numerous testimonies of the importance of hunting in the Mesoamerican ritual life101. While the deer was a prey of choice in different societies, associated with elite rituals and ceremonial hunting102, the capture or hunting of wolves could have taken place in the same context.
On the contrary, as a species of open and disturbed environment, the coyote may have thrived in past agricultural landscapes and it can be considered as a commensal species. Its formal identification in the archaeological record has proven to be problematic and often requires the use of ancient DNA103. Using a systematic GMM approach, we suggest that only a small minority of coyotes were present in the archaeological sites of central Mexico, which confirms previous zooarchaeological results. The scarcity of the coyote in the Mesoamerican archaeological record could be related to a cultural bias and a conscious choice of wolves against coyotes in ceremonial paraphernalia and state-level rituals, due to ecological and ethological distinctions. But whereas today coyotes proliferate in central Mexico they may have been only occasional in the past. The development of agropastoralism leading to land clearance and increasing hunting pressure on its natural predator, the wolf, has allowed the coyote to extend its natural distribution during the past 500 years104. Its previous natural distribution is not well understood and the mountain range of central Mexico could have represented its southernmost limit. This hypothesis would strengthen previous suggestions that the wild canids represented in the iconography, in particular in the site of Teotihuacan, are wolves rather than coyotes105.
Conclusion and future directions
In this paper, we used a GMM approach to identify first molars of large canids from 4 archaeological sites in central Mexico. We demonstrate that the dog, the wolf and the coyote can be differentiated using the size and shape of their teeth. Using the same approach on unidentified archaeological samples, we succeeded in the confident identification of 18 dogs, as well as two possible dogs and one possible coyote. One sample from the archaeological site of Tizayuca remains unidentified.
Overall, we confirm the prominent place of dogs in ancient Mesoamerica and the typical association of wolf remains and state-level rituals. The scarcity of coyote remains suggest this species was not as highly valued as the two others, whether it was in economic or symbolic terms. The ambiguous identification of some individuals that present an intermediate phenotype also led us to raise the question of the presence of hybrids, although a larger study would be required to fully address this question.
This preliminary study demonstrates the potential of a GMM approach for the identification of canid species in ancient Mesoamerica. However, it still relies on a very limited amount of reference individuals that will have to be expanded in the future. The confrontation of the GMM results to a genetic identification, through the analysis of ancient mitochondrial and nuclear DNA, would also be necessary to confirm the species identification and identify the degree of admixture between taxa.
Acknowledgments
We wish to thank Norma Valentín, Joaquín Arroyo Cabrales and Fabiola Guzmán for granting us access to the collection of the Laboratorio de Arqueozoología M. en C. Ticul Álvarez del Instituto Nacional de Antropología e Historia (Mexico city); Cécile Callou and Christine Lefèvre for facilitating the access to the Collections d’anatomie comparée of the Muséum national d’Histoire naturelle (Paris); Myriam Boudadi-Maligne and Jean-Baptiste Mallye for organising the conference Relations Hommes-canidés in Bordeaux in October 2018 and accepting our presentation; an anonymous reviewer for insightful comments.
Bibliography •••
- Adams, D. C., Collyer, M. L. and Kaliontzopoulou, A. (2019): Geomorph: Software for Geometric Morphometric Analyses. https://github.com/geomorphR/geomorph
- Adams, D. C., Rohlf, F. J. and Slice, D. E. (2004): “Geometric morphometrics: ten years of progress following the ‘Revolution’”, Italian Journal of Zoology, 71, 5-16.
- Adams, D. C., Rohlf, F. J. and Slice, D. E. (2013): “A field comes of age: geometric morphometrics in the 21st Century”, Hystrix, 24, 7-14.
- Adams, J. R., Leonard, J. and Waits, L. P. (2003): “Widespread occurrence of a domestic dog mitochondrial DNA haplotype in Southeastern US coyotes”, Molecular Ecology, 12, 541-546.
- Álvarez, T. and Ocaña, A. (1999): Sinopsis de los restos arqueozoológicos de vertebrados terrestres, México.
- Amano, N. (2011): The faunal remains of Nagsabaran in Cagayan, Northern Philippines: subsistence strategies in the Late Holocene. MSc thesis, University of the Philippines.
- Aranda, M. & Lopez-de-Buen, L. (1995): “Hábitos alimentarios del Coyote (Canis latrans) en la Sierra del Ajusco, México”. Acta Zoológica Mexicana (Nueva Serie) 65, 89-99.
- Arroyo Cabrales, J. & Carranza Castañeda, O. (2009): “Los canidos prehistóricos mexicanos antes de la llegada del perro”, Archaeobios, 3, 34-45.
- van Asch, B., Zhang, A., Oskarsson, M. C. R., Klütsch, C. F. C., Amorim, A. and Savolainen, P. (2013): “Pre-columbian origins of native American dog breeds, with only limited replacement by European dogs, confirmed by mtDNA analysis” Proceedings of the Royal Society B, 280, 1-9.
- Balme, J., O’Connor, S. and Fallon, S. (2018): “New dates on dingo bones from Madura Cave provide oldest firm evidence for arrival of the species in Australia”, Scientific Reports, 8, 9933.
- Baxter, I. L. & Hamilton-Dyer, S. (2003): “Foxy in furs? A note on evidence for the probable commercial exploitation of the red fox (Vulpes vulpes L.) and other fur bearing mammals in Saxo-Norman (10th -12th century AD) Hertford, Hertfordshire, U.K.”, Archaeofauna, 12, 87-94.
- Baylac, M. and Friess, M. (2005): “Fourier descriptors, procrustes superimposition and data dimensionality: an example of cranial shape analysis in modern human population”, in: Slice, D., dir., Modern Morphometrics in Physical Anthropology, Developments in Primatology: Progress and Prospects book series (DIPR), 142-165.
- Blanco, A., Rodríguez, B. and Valadez, R. (2007): “El lobo mexicano (Canis lupus baileyi) en el contexto cultural prehispánico: las fuentes escritas”, AMMVEPE, 18, 68-76.
- Blanco Padilla, A., Rodríguez Galicia, B. and Valadez Azúa, R. (2009): Estudio de los cánidos arqueológicos del México prehispánico, México.
- Bogan, M. A. and Mehlhop, P. (1983): “Systematic relationships of gray wolves (Canis lupus) in Southwestern North America” Occasional Papers of the Museum of Southwestern Biology, 1, 1-21.
- Bond, J. M. (1996): “Burnt offerings: animal bone in Anglo-Saxon cremations”, World Archaeology, 28, 76-88.
- Boudadi-Maligne, M. and Escarguel, G. (2014): “A biometric re-evaluation of recent claims for Early Upper Palaeolithic wolf domestication in Eurasia”, Journal of Archaeological Science, 45, 80-89.
- Boudadi-Maligne, M., Mallye, J.-B., Castel, J.-C., Ferrié, J.-G., Crepin, L., Kuntz, D., Vercoutère, C., Soulier, M.-C. and Costamagno, S. (2018): “Loups, chiens et sociétés du Paléolithique Supérieur”, in: Costamagno, S., Gourichon, L., Dupont, C., Dutour, O., Vialou, D. , dir., Animal symbolisé, animal exploité : du Paléolithique à la Protohistoire, Paris, Édition électronique du CTHS, Actes des congrès des sociétés historiques et scientifiques, 198-213. https://www.researchgate.net/publication/328281125_Loups_chiens_et_societes_du_Paleolithique_superieur
- Boudadi-Maligne, M., Mallye, J.-B., Langlais, M. and Barshay-Szmidt, C. (2012) : “Des restes de chiens magdaléniens à l’Abri du Morin (Gironde, France). Implications socioéconomiques d’une innovation zootechnique”, PALEO, 23, 39-54.
- Brown, D. E. 2002. The wolf of the Southwest: the making of an endangered species, Silver City.
- Byrd, B. F., Cornellas, A., Eerkens, J. W., Rosenthal, J. S., Carpenter, T. R., Leventhal, A. and Leonard, J. A. (2013): “The role of canids in ritual and domestic contexts: new ancient DNA insights from complex hunter–gatherer sites in prehistoric central California”, Journal of Archaeological Science, 40, 2176-2189.
- Carballo, D. M. (2007): “Implements of state power. Weaponery and martially themed obsidian production near the Moon Pyramid, Teotihuacan”, Ancient Mesoamerica, 18, 173–190.
- Castroviejo-Fisher, S., Skoglund, P., Valadez, R., Vilá, C. and Leonard, J. A. (2011): “Vanishing native American dog lineages”, BMC Evolutionary Biology, 11, 1-5.
- Ceballos, G. and Arroyo Cabrales, J. (2012): “Lista actualizada de los mamíferos de México”, Revista Mexicana de Mastozoología Nueva Época, 2, 27-80.
- Clutton-Brock, J. and Hammond, N. (1994): “Hot dogs: comestible canids in Preclassic Maya culture at Cuello, Belize”, Journal of Archaeological Science, 21, 819-826.
- Cucchi, T., Mohaseb, A., Peigné, S., Debue, K., Orlando, L. and Mashkour, M. (2017): “Detecting taxonomic and phylogenetic signals in equid cheek teeth: towards new palaeontological and archaeological proxies”, Royal Society Open Science, 4, 160997.
- Cucchi, T., Hulme-Beaman, A., Yuan, J. and Dobney, K. (2011): “Early Neolithic pig domestication at Jiahu, Henan province, China: clues from molar shape analyses using geometric morphometric approaches”, Journal of Archaeological Science, 38, 11-22.
- Cucchi, T., Kovács, Z. E., Berthon, R., Orth, A., Bonhomme, F., Evin, A., Siahsarvie, R., Darvish, J., Bakhshaliyev, V. and Marro, C. (2013): “On the trail of Neolithic mice and men towards Transcaucasia: zooarchaeological clues from Nakhchivan (Azerbaijan)”, Biological Journal of the Linnean Society, 108, 917-928.
- Day, J. S., Butterwick, K. and Pickering, R. B. (1996): “Archaeological interpretations of West Mexican ceramic art from the Late Preclassic period: three figurine projects”, Ancient Mesoamerica, 7, 149-161.
- Dehouve, D. (2010): “Un ritual de cacería. El conjuro para cazar venados de Ruiz de Alarcón”, Estudios de Cultura Nahuatl, 40, 299-331.
- Díaz del Castillo, B. (1996): La conquête du Mexique, Paris.
- Drake, A. G., Coquerelle, M., Kosintsev, P. A., Bachura, O. P., Sablin, M., Gusev, A. V., Lacey S. Fleming, L. S. and Losey, R. J. (2017). “Three-dimensional geometric morphometric analysis of fossil canid mandibles and skulls”, Scientific Reports, 7, 9508.
- Drake, A. G. and Klingenberg, C. P. (2010): “Large-scale diversification of skull shape in domestic dog: disparity and modularity”, The American Naturalist, 175, 289-301.
- Emery, K. F. (1999): “Continuity and variability in Postclassic and Colonial animal use at Lamanai and Tipu, Belize”, in: White, C.D., dir., Reconstructing Ancient Maya Diet, University of Utah Press, Salt Lake City, 61-83. https://www.academia.edu/873113/Continuity_and_variability_in_Postclassic_and_colonial_animal_use_at_Lamanai_and_Tipu_Belize
- Emery, K. F. (2004): “In search of the ‘Maya diet’: is regional comparison possible in the Maya tropics?”, Archaeofauna, 13, 37-55.
- Equihua Manrique, J. C., Tirado Balandrán, A. L., Ponce Ordaz, K. P., Nicolás Careta, C., Hernández Balderas, T., López Martínez, H., Padilla Gutiérrez, E. F., Riego Ruíz, S., Cervantes Alejandre, G. S., Montes Guerrero, T., López Martínez, J. B., Monterroso Rivas, P. N., Ponce de León, R. H. and Ojeda Güitró, E. (2008): Proyecto Salvamento Arqueológico Tizayuca 2006. Informe. Secuencias Estratigráficas y Conjuntos Arquitectónicos, Tomo I. Final Report, Instituto Nacional de Antropología e Historia.
- Evin, A., Cucchi, T., Cardini, A., Vidarsdottir, U. S., Larson, G. and Dobney, K. (2013): “The long and winding road: identifying pig domestication through molar size and shape”, Journal of Archaeological Science, 40, 735-743.
- Evin, A., Cucchi, T., Escarguel, G., Owen, J., Larson, G., Vidarsdottir, U. S. and Dobney, K. (2014): “Using traditional biometrical data to distinguish west palearctic wild boar and domestic pigs in the archaeological record: new methods and standards”, Journal of Archaeological Science, 43, 1-8.
- Evin, A., Dobney, K., Schafberg, R., Owen, J., Vidarsdottir, U. S., Larson, G. and Cucchi, T. (2015). “Phenotype and animal domestication: a study of dental variation between domestic, wild, captive, hybrid and insular Sus scrofa”, BMC Evolutionary Biology, 15, 1-16.
- Faugère, B. (2008): “Le cerf chez les anciens P’urhépecha du Michoacán (Mexique) : guerre, chasse et sacrifice”, Journal de la Société des Américanistes, 94, 109-142.
- Fillios, M. A. and Taçon, P. S. C. (2016): “Who let the dogs in? A review of the recent genetic evidence for the introduction of the dingo to Australia and implications for the movement of people”, Journal of Archaeological Science: Reports, 7, 782-792.
- Fisher, A. E. (2019): “When is a wolf a dog? combined geometric morphometrics and stable isotope analyses for differentiating wild from domestic canids on the Northern Plains”, Plains Anthropologist, 64, 316-349.
- Fritzell, E. K. and Haroldson, K. J. (1982): “Urocyon Cinereoargenteus”, Mammalian Species, 189, 1-8.
- García Moreno, J., Matocq, M. D., Roy, M. S., Geffen, E. and Wayne, R. K. (1996): “Relationships and genetic purity of the endangered Mexican wolf based on analysis of microsatellite loci”, Conservation Biology, 10, 376-389.
- Gentry, A., Clutton-Brock, J. and Groves, C. P. 2004. “The naming of wild animal species and their domestic derivatives”, Journal of Archaeological Science, 31, 645-651.
- Germonpré, M., Sablin, M. V., Stevens, R. E., Hedges, R. E. M., Hofreiter, M., Stiller, M. and Després, V. R. (2009): “Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes”, Journal of Archaeological Science, 36, 473-490.
- Gerry, J. P. (1997): “Bone isotope ratios and their bearing on elite privilege among the Classic Maya”, Geoarchaeology: An International Journal, 12, 41-69.
- Götz, C. M. (2008): “Coastal and inland patterns of faunal exploitation in the prehispanic northern Maya lowlands”, Quaternary International, 191, 154-169.
- Grandal-d’Anglade, A., Albizuri, S., Nieto, A., Majó, T., Agustí, B., Alonso, N., Antolín, F., López, J. B., Moya, A., Rodríguez, A. and Palomo, A. (2019): “Dogs and foxes in Early-Middle Bronze Age funerary structures in the Northeast of the Iberian peninsula: human control of canid diet at the sites of Can Roqueta (Barcelona) and Minferri (Lleida).” Archaeological and Anthropological Sciences, 11, 3949-3978.
- Guedes Milheira, R., Loponte, D. M., García Esponda, C., Acosta, A. and Ulguim, P. (2017): “The first record of a pre-Columbian domestic dog (Canis lupus familiaris) in Brazil”, International Journal of Osteoarchaeology, 27, 488- 494.
- Guthmann, E., Lefranc, P. and Arbogast, R.-M. (2016): “Un dépôt de renard roux (Vulpes vulpes) du 4e millénaire av. J.-C. à Entzheim ‘Les Terres de La Chapelle’ (Bas-Rhin) : offrande ou sépulture animale ?”, Revue Archéologique de l’Est, 65, 257-268.
- Hall, E. R. and Kelson, K. R. (1959): The Mammals of North America, vol. 2, New York.
- Hammer, Ø., Harper, D. A. T. and Ryan, P. D. (2001): “PAST: Paleontolgical statistics software package for education and data analysis”, Palaeontologica Electronica, 4, 1-9.
- Harding, L. E., Heffelfinger, J., Paetkau, D., Rubin, E., Dolphin, J. and Aoude, A. (2016): “Genetic management and setting recovery goals for Mexican wolves (Canis lupus baileyi) in the wild”, Biological Conservation, 203, 151-159.
- Hidalgo-Mihart, M. G., Cantu-Salazar, L., Gonzalez-Romero, A. and Lopez-Gonzalez, C. A. (2004): “Historical and present distribution of coyote (Canis latrans) in Mexico and Central America”, Journal of Biogeography, 31, 2025-2038.
- Hidalgo-Mihart, M. G., Cantu-Salazar, L., Lopez-Gonzalez, C. A., Martinez-Meyer, E. and González-Romero, A. (2001): “Coyote (Canis latrans) food habits in a tropical deciduous forest of Western Mexico”, The American Midland Naturalist, 146, 210-216.
- Hillson, S. (2005): Teeth, Cambridge.
- Hody, J. W. and Kays, R. (2018): “Mapping the expansion of coyotes (Canis latrans) across North and Central America”, ZooKeys, 759, 81-97.
- Hulme-Beaman, A., Claude, J., Chaval, Y., Evin, A., Morand, S., Vigne, J.-D., Dobney, K. and Cucchi, T. (2018): “Dental shape variation and phylogenetic signal in the Rattini tribe species of Mainland Southeast Asia”, Journal of Mammalian Evolution, 26, 435-446.
- Huster, A. C. and Smith, M. E. (2015): “A new archaeological chronology for Aztec-period Calixtlahuaca, Mexico”, Latin American Antiquity, 26, 3-25.
- Iljin, N. A. (1941): “Wolf-dog Genetics”, Journal of Genetics, 42, 359-414.
- Janssens, L.A.A. and Lawler, D.F. (2019): “Were foxes really domesticated, and were dogs really beasts of burden, during the Bronze Age in Northeast Spain?”, Archaeological and Anthropological Sciences, 11, 3995-3998.
- Jarquin P., A. M. and Martinez Vargas, E. (2004): “Ritos y Mitos Prehispanicos Nahuas En Dos Tumbas de La Campana, Colima”, Estudios de Cultura Náhuatl, 35, 75-87.
- Jernvall, J. and Thesleff, I. (2000): “Reiterative signalling and patterning during mammalian tooth morphogenesis”, Mechanisms of Development, 92, 19-29.
- Kemp, B. M., Judd, K., Monroe, C., Eerkens, J. W., Hilldorfer, L., Cordray, C., Schad, R., Reams, E., Ortman, S. G. and Kohler, T. A. (2017): “Prehistoric mitochondrial DNA of domesticate animals supports a 13th century exodus from the Northern US Southwest”, PLOS ONE, 12, e0178882.
- Kubler, G. (1972): “Jaguars in the valley of Mexico”, in: Benson, 19-44.
- Kupczik, K., Cagan, A., Brauer, S. and Fischer, M. S. (2017): “The dental phenotype of hairless dogs with FOXI3 haploinsufficiency”, Scientific Reports, 7, 54-59.
- Larson, G. and Bradley, D. G. (2014) “How much is that in dog years? The advent of canine population genomics”, PLoS Genetics, 10, e1004093.
- Larson, G., Karlsson, E. K., Perri, A., Webster, M. T., Ho, S. Y. W., Peters, J., Stahl, P. W., Piper, P. J., Lingaas, F., Fredholm, M., Comstock, K. E., Modiano, J. F., Schelling, C., Agoulnik, A. I., Leegwater, P. A., Dobney, K., Vigne, J.-D., Vilà, C., Andersson, L. and Lindblad-Toh, K. 2012. “Rethinking dog domestication by integrating genetics, archeology, and biogeography”, Proceedings of the National Academy of Sciences of the United States of America, 109, 8878-8883.
- Lawrence, B. (1967): “Early domestic dogs”, Zeitshrift Für Säugetierkunde, 32, 44-59.
- Lawrence, B. and Bossert, W. H. (1967): “Multiple character analysis of Canis lupus, latrans and familiaris, with a discussion of the relationships of Canis niger”, American Zoologist, 7, 223-232.
- Leonard, J. A., Wayne, R. A., Wheeler, J., Valadez Azúa, R., Guillén, S. and Vilá, C. (2002): “Ancient DNA evidence for Old World origin of New World dogs”, Science, 298, 1613-1616.
- Leopold, A. S. (1959): Wildlife of Mexico, the game birds and mammals, Berkley.
- López Austin, A. and López Luján, L. (2012) : Le Passé Indigène. Histoire Pré-coloniale Du Mexique, Paris.
- López Luján, L., Chávez Balderas, X., Zuñiga-Arellano, B., Aguirre Molina, A. and Valentín, N. (2012): “Un portal al inframundo. Ofrendas de animales sepultadas al pie del Templo Mayor de Tenochtitlan”, Estudios de Cultura Náhuatl, 44, 9-40.
- López Luján, L. and Chávez Balderas, X. (2010): “Al pie del Templo Mayor: excavaciones en busca de los soberanos mexicas”, in: López Luján, L. & McEwan, C., dir., Moctezuma II. Tiempo y destino de un governante, Instituto nacional de antropología e historia, 294-303. http://mesoweb.org/es/articulos/sub/Al-pie-del-Templo-Mayor.pdf
- Losey, R. J., Bazaliiskii, V. I., Garvie-Lok, S., Germonpré, M., Leonard, J. A., Allen, A. L., Katzenberg, M. A. and Sablin, M. V. (2011): “Canids as persons: Early Neolithic dog and wolf burials, Cis-Baikal, Siberia”, Journal of Anthropological Archaeology, 30,174-189.
- Lucas, S. G., Alvarado, G. E. and Vega, E. (1997): “The Pleistocene mammals of Costa Rica”, The Society of Vertebrate Palaeontology, 17, 413-427.
- Lyman, R. L. (2008): Quantitative Paleozoology, Cambrige.
- Manin, A. (2015): “Utilisation matérielle et symbolique des animaux dans le nord de la Mésoamérique entre le Classique et la Conquête (200-1521 apr. J.-C.)”, PhD thesis, Muséum national d’Histoire naturelle.
- Manin, A. (2017a): “Archéozoologie, géochimie et concepts de domestication en Mésoamérique – Zooarchaeology, geochemistry, and concepts of domestication in Mesoamerica”, Annales de La Fondation Fyssen, 32, 141-154.
- Manin, A. (2017b): “Estudio de los restos de fauna procedentes del sitio de Calixtlahuaca, Estado de México, México”, Technical report, Proyecto Arqueológico Calixtlahuaca.
- Manin, A. (in prep.): “Caza y cría de animales en el valle de Acámbaro a partir desl estudio de la fauna vertebrada de El Mezquital – Los Azules”, in: Faugère.
- Manin, A. and Lefèvre, C. (2016): “The use of animals in Northern Mesoamerica, between the Classic and the Conquest (200-1521 AD). An attempt at regional synthesis on central Mexico”, Anthropozoologica, 51, 127-147.
- Manin, A. and Lefèvre, C. (2018): “Utilización material y simbólica de los animales en Vista Hermosa”, in: Stresser-Péan, C., Vista Hermosa. Nobles, artesanos y mercaderes en los confines del mundo huesteco. Volumen III: El arte y la vida en la Huesteca posclásica, 3, 9-50.
- Manin, A., Ollivier, M., Bastian, F., Zazzo, A., Tombret, O., Equihua Manrique, J. C. and Lefèvre, C. (2018): “Can we identify the Mexican hairless dog in the archaeological record? Morphological and genetic insights from Tizayuca, Basin of Mexico”, Journal of Archaeological Science, 98, 128-136.
- Martinez de Leon Marmol, B. L. and Reyes Carlo, I. (2007) “Los perros como indicador estacional en los enterramientos humanos prehispánicos. Un caso de estudio en el valle de Zapotitlan, Puebla”, Estudios de Antropología Biológica, 13, 347-360.
- Matos Moctezuma, E. (1994): “Mesoamérica”, in: Manzanilla & López Luján, vol. 1, 95-119.
- Merkle, J. A., Krausman, P. R., Stark, D. W., Oakleaf, J. K. and Ballard, W. B. (2009): “Summer diet of the Mexican gray wolf (Canis lupus baileyi)”, The Southwestern Naturalist, 54, 480-524.
- Monagle, V., Conrad, C. and Jones, E. L. (2018): “What makes a dog? Stable isotope analysis and human-canid relationships at Arroyo Hondo Pueblo”, Open Quaternary, 4, 6.
- Morey, D. F. (2006): “Burying key evidence: the social bond between dogs and people”, Journal of Archaeological Science, 33, 158-175.
- Morey, D. F. (2014): “In search of Paleolithic dogs: a quest with mixed results”, Journal of Archaeological Science, 52, 300-307.
- Morey, D. F. and Jeger, R. (2017): “From wolf to dog: Late Pleistocene ecological dynamics, altered trophic strategies, and shifting human perceptions”, Historical Biology, 29, 895-903.
- de Moura Bubadué, J., Cáceres, N., dos Santos Carvalho, R. and Meloro, C. (2016): “Ecogeographical variation in skull shape of South-American canids: abiotic or biotic processes?”, Evolutionary Biology, 43, 145-159.
- Napierala, H. and Uerpmann, H.-P. (2012): “A ‘new’ Palaeolithic dog from Central Europe”, International Journal of Osteoarchaeology, 22, 127-137.
- Ní Leathlobhair, M., Perri, A. R., Irving-Pease, E. K., Witt, K. E., Linderholm, A., Haile, J., Lebrasseur, O., Ameen, C., Blick, J., Boyko, A. R., Brace, S., Nunes Cortes, Y., Crockford, S. J., Devault, A., Dimopoulos, E. A., Eldridge, M., Enk, J., Gopalakrishnan, S., Gori, K., Grimes, V., Guiry, E., Hansen, A. J., Hulme-Beaman, A., Johnson, J., Kitchen, A., Kasparov, A. K., Kwon, Y.-M., Nikolskiy, P. A., Peraza Lope, C., Manin, A., Martin, T., Meyer, M., Myers, K. N., Omura, M., Rouillard, J.-M., Pavlova, E. Y., Sciulli, P., Sinding, M.-H. S., Strakova, A., Ivanova, V. V., Widga, C., Willerslev, E., Pitulko, V. V., Barnes, B., Gilbert, M. T. P., Dobney, K. M., Malhi, R. S., Murchison, E. P., Larson, G. and Frantz, L. A. F. (2018): “The evolutionary history of dogs in the Americas”, Science, 361, 81-85.
- Olivier, G. (2015): Caceria, sacrificio y poder en Mesoamérica. Tras las huellas de Mixcoatl, “Serpiente de Nube”, México.
- Olsen, S. J. (1985): Origins of the domestic dog, Tuscon.
- Ovodov, N. D., Crockford, S. J., Kuzmin, Y. V., Higham, T. F. G., Hodgins, G. W. L. and van der Plicht, J. (2011): “A 33,000-year-old incipient dog from the Altai mountains of Siberia: evidence of the earliest domestication disrupted by the Last Glacial Maximum”, PLoS ONE, 6, e22821.
- Pereira, G. (2008) : “L’archéologie de la Mésoamérique Septentrionale vue depuis le massif de Barajas (Guanajuato)”, Les Nouvelles de l’archéologie, 111/112, 101-106.
- Perri, A. (2016) : “A Wolf in dog’s clothing: initial dog domestication and Pleistocene wolf variation”, Journal of Archaeological Science, 68, 1-4.
- Pionnier-Capitan, M. (2010): La domestication du chien en Eurasie: étude de la diversité passée, approches ostéoarchéologiques, morphométriques et paléogénétiques, PhD thesis, École Normale Supérieure de Lyon.
- Pionnier-Capitan, M., Bemilli, C., Bodu, P., Célérier, G., Ferrié, J.-G., Fosse, P., Garcià, M. and Vigne, J.-D. (2011): “New evidence for Upper Palaeolithic small domestic dogs in South-Western Europe”, Journal of Archaeological Science, 38, 2123-2140.
- Pluskowski, A. (2006): “Where are the wolves? Investigating the scarcity of European grey wolf (Canis lupus lupus) remains in Medieval archaeological contexts and its implications”, International Journal of Osteoarchaeology, 16, 279-295.
- Porter, M. (1956): “Excavations at Chupicuaro, Guanajuato, Mexico”, Transactions of the American Philosiphical Society, 46, 515-637.
- Prates, L., Prevosti, F. J. and Berón, M. (2010): “First records of Prehispanic dogs in southern South America (Pampa-Patagonia, Argentina)”, Current Anthropology, 51, 273-280.
- <a href= »https://www.R-project.org/« target= »_blank »>R Core Team (2019): R: A language and environment for statistical computing.</a>
- Reed, J. E., Ballard, W. B., Gipson, P. S., Kelly, B. T., Krausman, P. R., Wallace, M. C. and Wester, D. B. (2006): “Diets of free‐ranging Mexican gray wolves in Arizona and New Mexico”, Wildlife Society Bulletin, 34, 1127-1133.
- Renaud, S., Alibert, P. and Auffray, J.-C. (2009): “Mandible shape in hybrid mice”, Naturwissenschaften, 96, 1043-1050.
- Rodríguez Galicia, B., Valadez Azúa, R., Pereira, G., Viniegra Rodríguez, F., Olmos Rodríguez, K. and Blanco Padilla, A. (2001): “Restos arqueozoológicos de perros (Canis familiaris) encontrados en el sitio de Guadalupe, estado de Michoacán”, AMMVEPE, 12, 199-209.
- <a href= »https://life.bio.sunysb.edu/morph/« target= »_blank »>Rohlf, F. J. (2015): “TpsDig, digitize landmarks and outlines, Version 2.19”.</a>
- Russell, N. (2002): “The wild side of animal domestication”, Society & Animals, 10, 285-302.
- de Sahagun, B. (1829): Historia general de las cosas de Nueva España vol. 1, México.
- de Sahagun, B. (1830): Historia general de las cosas de Nueva España vol. 2, México.
- Schlager, S. (2017): “Morpho and Rvcg – Shape Analysis in R”, in: Zheng, Li and Szekely, 217-256.
- Schmitt, E. and Wallace, S. (2014): “Shape change and variation in the cranial morphology of wild canids (Canis lupus, Canis latrans, Canis rufus) compared to domestic dogs (Canis familiaris) using geometric morphometrics”, International Journal of Osteoarchaeology, 24, 42-50.
- Schmutz, S. M., Berryere, T. G., Barta, J. L., Reddick, K. D. and Schmutz, J. K. (2007): “Agouti sequence polmorphisms in coyote, wolves and dogs suggest hybridization”, Journal of Heredity, 98, 351-355.
- Seetah, K., Cucchi, T., Dobney, K. and Barker, G. (2014): “A geometric morphometric re-evaluation of the use of dental form to explore differences in horse (Equus caballus) populations and its potential zooarchaeological application”, Journal of Archaeological Science, 41, 904-910.
- Seler, E. (1996): “The animal pictures of the Mexican and Mayan manuscripts”, in: Collected works in Mesoamerican linguistics and archaeology vol. 5, 165-340.
- Shirokova, V., Jussila, M., Hytönen, M. K., Perälä, N., Drögemüller, C., Leeb, T., Lohi, H., Sainio, K., Thesleff, I. and Mikkola, M. L. (2013): “Expression of Foxi3 is regulated by ectodysplasin in skin appendage placodes”, Developmental Dynamics, 242, 593-603.
- Sugiyama, N. (2014): Animals and sacred mountains: how ritualized performances materialized state-ideologies at Teotihuacan, Mexico. PhD thesis, Harvard University.
- Sugiyama, N., Pérez, G., Rodríguez, B., Torres, F. and Valadez, R. (2014) : “The role of animals in state-level rituals in Mesoamerica”, in: Arbuckle, B. & McCarty, S.A., dir., Animals and Inequality in the Ancient World, University Press of Colorado, 11-31.
- Sugiyama, S. and López Luján, L. (2006): “Simbolismo y función de los entierros dedicatorios de la Pirámide de la Luna en Teotihuacan”, in: Lopez Luján, L., Carrasco, D., Cué, L., dir., Arqueología e historia del Centro de México. Homenaje a Eduardo Matos Moctezuma, Instituto nacional de antropología e historia, 131-151.
- Thesleff, I. (2006): “The genetic basis of tooth development and dental defects”, American Journal of Medical Genetics Part A, 14, 2530-2535.
- Tomaszewski, B. M. and Smith, M. E. (2011): “Polities, territory and historical change in Postclassic Matlatzinco (Toluca Valley, Central Mexico)”, Journal of Historical Geography, 37, 22-39.
- Valadez Azúa, R., Blanco Padilla, A., Rodríguez Galicia, B. and Pérez Roldán, G. (2013): “The dog in the Mexican archaeozoological record”, in: Götz, C. M. & Emery, K. F., dir., The Archaeology of Mesoamerican Animals, Lockwood press, 557-582. https://www.researchgate.net/publication/304822224_The_Dog_in_the_Mexican_Archaeozoological_Record
- Valadez Azúa, R. & Rodríguez Galicia, B. (2014): “Uso de la fauna, estudios arqueozoológicos y tendencias alimentarias en culturas prehispánicas del centro de México”, Anales de Antropología, 48, 139-166.
- Valadez Azúa, R. and Rodríguez Galicia, B. (2009): “Canidos presentes en el proyecto ‘Tuneles y cuevas de Teotihuacan’”, in: Manzanilla Naim, L. R., dir., Obras 1, El inframundo de Teotihuacan. Ocupaciones post-teotihuacanas en los túnels al este de la pirámide del sol, vol. II, El ambiente y el hombre, El colegio nacional, 573-669. http://www.iia.unam.mx/directorio/archivos/MANL510125/2009_Manzanilla-El-Inframundo-de-Teotihuacan_Fauna.pdf
- Valadez Azúa, R., Rodríguez Galicia, B., Manzanilla, L. and Tejeda, S. (2006): “Dog-wolf hybrid biotype reconstruction from the archaeological city of Teotihuacan in Prehispanic central Mexico”, in: Snyder, L. M. & Moore, E. A., dir., Dogs and People in Social, Working, Economic or Symbolic Interaction, 9th ICAZ Conference, Durham, 2002, 121-131. https://www.researchgate.net/publication/304824785_Dog-wolf_Hybrid_Biotype_Reconstruction_from_the_Archaeological_City_of_Teotihuacan_in_Prehispanic_Central_Mexico
- Valentín Maldonado, N. (1997): “Análisis del material zoológico procedente de los sitios arqueológicos de Jonuta, Tabasco y Campeche, México”, in: Arroyo Cabrales, J. & Polaco, J., dir., Homenaje al profesor Ticul Álvarez, Colección Científica, 357, México, 343-366.
- Valenzuela-Lamas, S., Baylac, M., Cucchi, T. and Vigne, J.-D. (2011): “House mouse dispersal in Iron Age spain: a geometric morphometrics appraisal”, Biological Journal of the Linnean Society, 102, 483-497.
- Vigne, J.-D. (2011): “The origins of animal domestication and husbandry: a major change in the history of humanity and the biosphere”, Comptes Rendus Biologies 334, 171-181.
- vonHoldt, B. M., Cahill, J. A., Fan, Z., Gronau, I., Robinson, J., Pollinger, J. P., Shapiro, B., Wall, J. and Wayne, R. K. (2016a): “Whole-genome sequence analysis shows that two endemic species of North American wolf are admixtures of the coyote and gray wolf”, Science Advances, 2, e1501714.
- vonHoldt, B. M., Kays, R., Pollinger, J. P. and Wayne, R. K. (2016b): “Admixture mapping identifies introgressed genomic regions in North American canids”, Molecular Ecology, 25, 2443-2453.
- vonHoldt, B. M., Pollinger, J. P., Earl, D. A., Knowles, J. C., Boyko, A. R., Parker, H., Geffen, E., Pilot, M., Jedrzejewski, W., Jedrzejewska, B., Sidorovich, V., Greco, C., Randi, E., Musiani, M., Kays, R., Bustamante, C. D., Ostrander, E. A., Novembre, J. and Wayne, R. K. (2011): “A genome-wide perspective on the evolutionary history of enigmatic wolf-like canids”, Genome Research, 21, 1294-1305.
- Wayne, R. K., Lehman, N., Allard, M. W. and Honeycutt, R. L. (1992): “Mitochondrial DNA variability of the gray wolf: genetic consequences of population decline and habitat fragmentation”, Conservation Biology, 6, 559-569.
- Walker, D. N. and Frison, G. C. (1982): “Studies on Amerindian dogs, 3: Prehistoric wolf/dog hybrids from the Northwestern plains”, Journal of Archaeological Science, 9, 125-172.
- White, C. D. (2004): “Stable isotopes and the human-animal interface in Maya biosocial and environmental systems”, Archaeofauna, 13, 183-198.
- White, C. D., Pohl, M. E. D. D., Schwarcz, H. P. and Longstaffe, F. J. (2001): “Isotopic evidence for Maya patterns of deer and dog use at Preclassic Colha”, Journal of Archaeological Science, 28, 89-107.
- White, C. D. and Schwarcz, H. P. (1989): “Ancient Maya diet as inferred from isotopic and elemental analysis of human bone”, Journal of Archaeological Science, 16, 451-474.
- White, C. D., Pohl, M. E. D. D., Schwarcz, H. P. and Longstaffe, F. J. (2004): “Feast, field and forest: Deer and dog diets at Lagartero, Tikal, and Copán”, in: Emery, K. F., ed., Mayan zoorchaeolgy: new directions in method and theory, Monograph, vol. 51, 141-158.
- Wilson, D. E. and Reeder, D. A. E. (2005): Mammals Species of the World, 3rd edition, Baltimore.
- Zeder, M. A., Emshwiller, E., Smith, B. D. and Bradley, D. G. (2006): “Documenting domestication: the intersection of genetics and archaeology”, Trends in Genetics, 22, 139-155.
Notes
- e.g. Russell 2002; Vigne 2011; Zeder et al. 2006.
- In this chapter, we chose to use the nomenclature suggested by Gentry et al. (2004) and refer to the dog as Canis familiaris, and not Canis lupus familiaris as advised by the taxonomic authorities (e.g. the Integrative Taxonomic Information System, http://www.itis.gov), in order to highlight the difference of status between the dog and the wolf.
- e.g. Boudadi-Maligne & Escarguel 2014; Germonpré et al. 2009; Larson & Bradley 2014; Morey 2014; Ovodov et al. 2011; Perri 2016.
- Boudadi-Maligne et al. 2012; Napierala & Uerpmann 2012; Pionnier-Capitan et al. 2011.
- Boudadi-Maligne et al., 2018; Larson et al. 2012; Morey 2006; Morey & Jeger 2017.
- Balme et al. 2018; Fillios & Taçon 2016.
- Guedes Milheira et al. 2017; Prates et al. 2010.
- Grandal-d’Anglade et al. 2019 (but see Janssens & Lawler 2019 for a more nuanced interpretation); Guthmann et al., 2016.
- Baxter & Hamilton-Dyer 2003; Bond 1996.
- e.g. Losey et al. 2011; Pluskowski 2006; Sugiyama et al. 2014.
- e.g. Lawrence 1967; Olsen 1985; Pluskowski 2006; Walker & Frison 1982.
- The status of two taxa, the Eastern wolf (Canis lycaon) and the red wolf (Canis rufus), is still debated as they arise from a fairly recent admixture between the wolf (Canis lupus) and the coyote (Canis latrans) while presenting increased fitness to their environment (vonHoldt et al. 2016a; 2016b).
- Wilson & Reeder 2005.
- Mesoamerica is a cultural area ranging from central Mexico to western Honduras arising among agricultural societies ca. 2500 a.C. Despite encompassing regions of contrasting environments, constant exchanges and trade allowed the emergence of a common tradition and its consolidation up to the beginning of the 16th century p.C. (López Austin & López Luján 2012; Matos Moctezuma 1994).
- Arroyo Cabrales & Carranza Castañeda 2009; Ceballos & Arroyo Cabrales 2012.
- Fritzell & Haroldson 1982.
- e.g. Alvarez & Ocaña 1999; Götz 2008.
- Seler 1996, 193.
- Leonard et al. 2002; Ní Leathlobhair et al. 2018.
- Ameen et al. 2019; Ní Leathlobhair et al. 2018.
- van Asch et al. 2013; Castroviejo-Fisher et al. 2011; Ní Leathlobhair et al. 2018.
- Manin 2017a; White 2004.
- Gerry 1997; White et al. 2001; White & Schwarcz 1989.
- White 2004; White et al. 2004.
- Bogan & Mehlhop 1983; Hall & Kelson 1959 vol. 2, 847.
- García Moreno et al. 1996; von Holdt et al. 2011; Wayne et al. 1992.
- Leopold 1959, 401.
- Brown 2002; cited by Harding et al. 2016.
- Harding et al. 2016.
- Leopold 1959, 401.
- Merkle et al. 2009; Reed et al. 2006.
- Lucas et al. 1997.
- Hody & Kays 2018.
- Hidalgo-Mihart et al. 2004; Hody & Kays 2018.
- Aranda & Lopez-de-Buen 1995; Hidalgo-Mihart et al. 2001.
- Adams et al. 2003; Schmutz et al. 2007.
- vonHoldt et al. 2011; 2016a.
- Ní Leathlobhair et al. 2018.
- de Sahagun 1829, 263.
- Martínez de León Mármol & Reyes Carlo 2007.
- Jarquin P. & Martinez Vargas 2004.
- Rodríguez Galicia et al. 2001.
- Seler 1996, 182.
- Day et al. 1996.
- Diaz del Castillo 1996, 334; de Sahagun 1829, 160.
- e.g. Clutton-Brock & Hammond 1994; Emery 2004; Manin & Lefèvre 2018; Valadez Azúa et al. 2013.
- de Sahagun 1830, 524.
- Blanco et al. 2007.
- e.g. Carballo 2007; Kubler 1972; Seler 1996, 193.
- Valadez Azúa & Rodríguez Galicia 2009, 622.
- Sugiyama et al. 2014.
- Manin 2015, 316.
- Emery 1999.
- e.g. Álvarez & Ocaña 1999; Valentín Maldonado 1997.
- Sugiyama & López Luján 2006; Sugiyama et al. 2014.
- Álvarez & Ocaña 1999, 82.
- Álvarez & Ocaña, 1999, 82.
- López Luján & Chavez Balderas 2010; López Luján et al. 2012.
- Álvarez & Ocaña 1999, 82.
- Valentín Maldonado, com. pers. 2014.
- Adams et al. 2004, 2013.
- Jernvall & Thesleff 2000; Thesleff 2006.
- Hillson 2005, 152.
- Cucchi et al. 2013; Hulme-Beaman et al. 2018; Valenzuela-Lamas et al. 2011.
- Cucchi et al. 2011; Evin et al. 2013; 2015.
- Cucchi et al. 2017; Seetah et al. 2014.
- Ameen et al. 2019; Drake et al. 2017; Drake & Klingenberg 2010; Fisher 2019; de Moura Bubadué et al. 2016; Schmitt & Wallace 2014.
- Amano 2011; Pionnier-Capitan 2010.
- Manin et al. 2018; Ní Leathlobhair et al. 2018.
- Our study was carried out mainly in the collections of Anatomie Comparée of the Muséum national d’Histoire naturelle, with a short complementary stay in the Laboratorio de Arqueozoología M. en C. Ticul Álvarez del Instituto Nacional de Antropología e Historia, Mexico.
- Lawrence & Bossert 1967.
- Iljin 1941.
- vonHoldt et al. 2011.
- Blanco Padilla et al. 2009.
- Kupczik et al. 2017; Shirokova et al. 2013.
- Manin et al. 2018.
- Manin et al. 2018; Ní Leathlobhair et al. 2018.
- Pereira 2008.
- Porter, 1956.
- Faugère in prep.
- Manin in prep.
- Huster & Smith 2015; Tomaszewski & Smith 2011.
- Manin 2017b; 2015, 160.
- Equihua et al. 2008.
- Manin et al. 2018; Ní Leathlobhair et al. 2018.
- Rohlf 2015.
- R Core Team 2019.
- Schlager 2017.
- Baylac & Friess 2005.
- R Core Team, 2019.
- Manin & Lefèvre 2016; Valadez Azúa & Rodríguez Galicia 2014.
- Manin et al. 2018.
- Lyman 2008, 180.
- López Luján & Chávez Balderas 2010; Lopez Lujan et al. 2012; Sugiyama et al. 2014; Valadez Azúa & Rodriguez Galicia 2009.
- ibid.
- Ní Leathlobhair et al. 2018.
- Adams et al. 2003; Schmutz et al. 2007 vonHoldt et al. 2011; 2016b.
- Valadez Azúa et al. 2006; Walker & Frison 1982.
- e.g. Evin et al. 2015.
- e.g. Renaud et al. 2009.
- e.g. Olivier 2015.
- Dehouve 2010; Faugère 2008; Olivier 2015.
- Byrd et al. 2013; Kemp et al. 2017; Monagale et al. 2018.
- Hidalgo-Mihart et al. 2004; Hody & Kays 2018.
- Sugiyama 2014.
