




Introduction
Au Pléistocène en Europe, les études paléontologiques ont permis d’appréhender avec précision l’évolution des Canidés, en particulier du genre Canis1. En parallèle, différents rôles joués leur sont reconnus pour les périodes du Paléolithique et de l’Épipaléolithique d’Europe et du Levant : accumulateurs dans les cavités, en particulier pour le genre Cuon2 ; chapardeurs3 ; proies consommées ou utilisées pour la pelleterie4 ; et enfin animaux domestiqués dès le Paléolithique supérieur en Europe et l’Épipaléolithique au Levant, bien qu’il n’y ait toujours aucun consensus sur la période d’apparition de la domestication du chien5.
En revanche, en Afrique du Nord-Ouest, les études sur l’évolution et la place des Canidés dans les assemblages fossiles sont plus rares6. En effet, les faunes modernes se sont mises en place progressivement au cours du Pléistocène. Entre 8 et 2 Ma la présence du genre Nyctereutes est mentionnée, avec notamment l’espèce Nyctereutes abdeslami à Ahl al Oughlam au Maroc. Pour les renards, entre 4 Ma et l’actuel se développe le genre Vulpes, avec Vulpes hassani reconnu dès la fin du Pliocène à Ahl al Oughlam, suivi de Vulpes cf. rueppellii à Tighennif à la fin du Pléistocène inférieur en Algérie7. À l’Aïn Boucherit (Algérie), que Sahnouni et al.8 datent d’environ 2,6 Ma (voir aussi9), on note la présence d’un Vulpes sp.10, une mention rejetée par Geraads11. L’espèce Vulpes rueppellii est ensuite à nouveau mentionnée dans le Pléistocène supérieur de la grotte des Contrebandiers au Maroc12. L’espèce Vulpes vulpes ne semble arriver qu’au Pléistocène supérieur, dans des sites comme Doukkala ou El Harhoura 1, pour perdurer à l’Holocène. Notons également, plus rare, l’espèce Vulpes pallida13. Enfin, il faut souligner la mention d’un reste de Vulpes zerda à la grotte des Contrebandiers14. Actuellement, certaines de ces formes de Renard sont encore présentes en Afrique du Nord, dont le Renard famélique, Vulpes rueppellii, le Renard roux, Vulpes vulpes et le Fennec Vulpes zerda15. Pour les autres Canidés, de 8 Ma à l’actuel sont mentionnées plusieurs espèces du genre Lupulella : Lupulella paralius à Ahl al Oughlam et Lupulella mohibi à la Grotte à Hominidés (Maroc)16. Dans le genre Canis, l’espèce Canis primaevus a été citée dans les sites d’Aïn Boucherit et d’Aïn Kerba17. À Aïn Hanech (Algérie), D. Geraads18 parle de Canis africanus (anciennement nommé Xenocyon lycaonoides), tandis que Sahnouni et al.19 parlent de Canis cf. atrox, mais quoiqu’il en soit il s’agit d’une forme voisine du loup. À l’Oued Sarrat (Tunisie), c’est une forme originale, peut-être voisine de Lupulella mohibi, Canis othmanii qui est présente à 1 Ma environ20.
L’espèce Canis aureus a été citée dans de nombreux niveaux du Pléistocène supérieur et de l’Holocène, mais des analyses génétiques suggèrent que le grand Chacal égyptien actuel, Canis aureus lupaster, se rapprocherait plus du Loup (Canis lupus) que du Chacal européen (Canis aureus) et pourrait donc être attribué à Canis lupus lupaster ou à Canis anthus, ce qui remet en question l’existence du Chacal eurasiatique en Afrique du Nord21. Canis lupus lupaster était-il déjà présent au Pléistocène dans cette région de l’Afrique ? Et si oui, pourrait-il correspondre aux Canidés fossiles de grande stature jusqu’à maintenant attribués à Canis aureus ? Enfin, concernant le genre Lycaon, l’espèce Lycaon magnus décrite à Tighennif22 est remplacée vers 500 ka par Lycaon pictus, reconnu dans le remplissage du Pléistocène supérieur de Doukkala 2 (Fig. 1, 2)23.
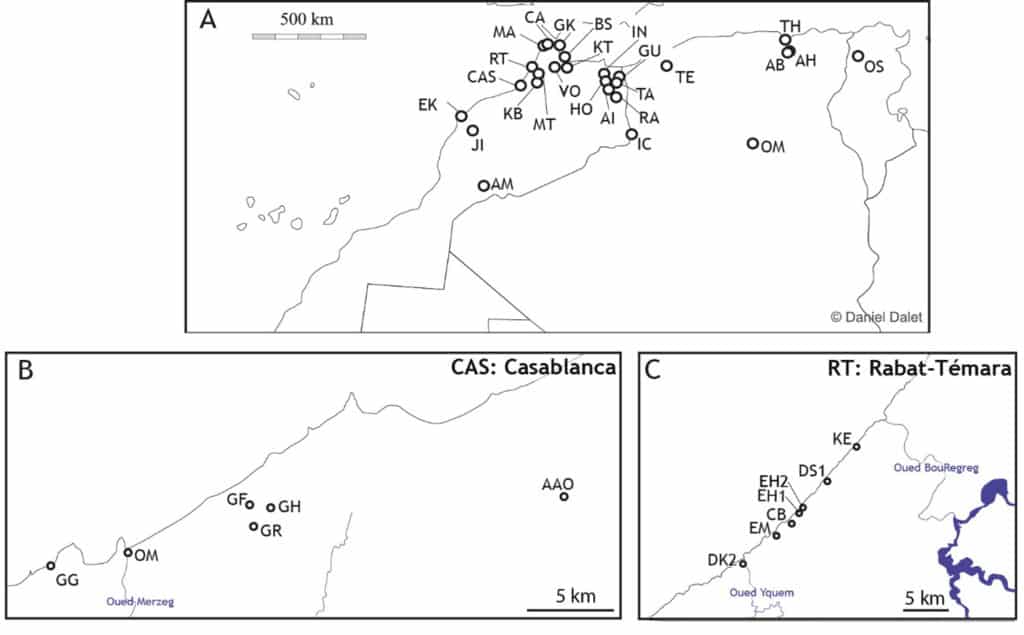
[AM : Adrar Metgourine, JI : Jebel Irhoud, EK : El Khenzira, CAS : Casablanca, KB : Kef-el-Baroud, RT : Rabat-Témara, MA : Mugharet el Aliya, CA : Sites du Cap Achakar, GK : Ghar-Khal, BS : Bou Saria, MT : M’Tsogatin 1, VO : Volubilis, KT : Kaf-Taht-el-Ghar, IN : Ifri n’Ammar, HO : Hassi Oenzouga, GU : Guenfouda, TA : Taforalt, AI : Aïn Metlili, RA : Rhafas, IC : Iche, TH : Tamar Hat ; AH : Aïn Hanech, AB : Aïn Boucherit, El-Kherba, TE : Ternifine, OM : Oued Metlili, OS : Oued Sarrat, GG : Grotte des Gazelles, OM : Oued Merzeg, GF : Grotte des Félins, GH : Grotte à Hominidés, GR : Grotte des Rhinocéros, AAO : Ahl al Oughlam, DK2 : Doukkala 2, EM : Grotte d’El Mnasra, CB : Grotte des Contrebandiers, EH1 : Grotte d’El Harhoura 1, EH2 : Grotte d’El Harhoura 2, DS1 : Grotte de Dar es Soltane 1, KE : Khebibat]
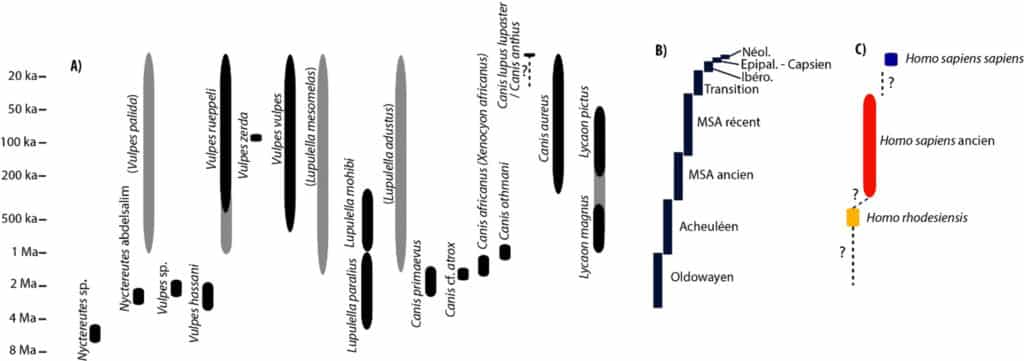
a. D’après Geraads (2011), Sahnouni et al. (2013), Koepfli et al. (2015), Amri et al. (2017),
Yuko Hallett (2018), des cultures ; b. Et des Hominidés ; c. Adapté de Hublin (2009).
À diverses époques, tous ces Canidés cohabitent ou ont cohabité avec de nombreux autres Carnivores : Hyénidés (Chasmaporthetes sp., Pliocrocuta sp., Hyaena hyaena, Crocuta crocuta), grands Félidés (Homotherium sp., Panthera leo, Panthera pardus, Acinonyx jubatus), Félidés de tailles moyenne et petite (Lynx thomasi, Caracal caracal, Leptailurus serval, Felis cf. silvestris, Felis lybica, Felis margarita), Ursidés (Ursus bibersoni, Ursus arctos), Mustélidés (Mellivora capensis, Lutra sp., Genetta genetta, Mustela nivalis, Ictonyx lybica), Viverridés (Viverra sp., Genetta sp.) et Herpestidés (Ichneumia sp., Herpestes ichneumon)24.
En parallèle, différentes espèces d’Hominidés se sont également succédées dans cette zone géographique, ainsi que différentes cultures :
- Le Early Stone Age, comprenant des outils oldowayens, est pour le moment reconnu seulement pour les gisements du Pléistocène inférieur d’Algérie tels que Aïn Boucherit25 ou Aïn Hanech26. Dès le début du Pléistocène moyen, des industries acheuléennes sont présentes dans les sites de Tighennif en Algérie et dans les carrières Thomas, Oulad Hamida et Sidi Abderrahmane à Casablanca au Maroc. Ces sites ont livré des restes humains attribués à Homo cf. mauritanicus / H. cf. rhodesiensis27.
- Le Middle Stone Age, avec le Middle Stone Age ancien (parfois dénommé Moustérien maghrébin) à partir de 310 ka associé à des restes d’Homo sapiens anciens présents dans le site de Jebel Irhoud28, suivi par le Middle Stone Age récent (parfois dénommé Atérien), daté entre 130 et 40/30 ka. Cette culture est associée à des restes humains attribués à des Homo sapiens anciens, avec comme sites emblématiques les grottes de Témara, de Taforalt, du Rhafas ou d’Ifri n’Ammar. Pour ces périodes, les industries peuvent comprendre des pièces pédonculées, des ornements, principalement confectionnés sur coquilles de gastéropodes marins (Nassariidae), de l’industrie osseuse et des blocs de colorant29.
- Une industrie de transition locale est décrite seulement au sein de la séquence stratigraphique de Taforalt, caractérisée par des pièces bifaciales originales30.
- L’Ibéromaurusien, correspondant au Paléolithique supérieur et au début de l’Épipaléolithique, qui se caractérise par la production de lamelles. Cette culture est datée entre 22 et 12 ka environ. Elle semble se scinder en deux phases autour de 15 ka. La majorité de la documentation sur ce faciès provient de la phase récente. Les industries sont associées à des restes d’Homo sapiens sapiens, parfois découverts en contexte de sépultures31. Une analyse génétique récente réalisée dans le site de Taforalt suggère une origine sub-saharienne de ces populations32.
- L’Épipaléolithique maghrébin et le Capsien, suivis par le Néolithique (Fig. 1, 2)33.
L’Afrique du Nord-Ouest est donc riche en restes de Canidés appartenant à diverses espèces. Néanmoins peu d’études s’intéressent à leurs rôles dans les sites. Ont-ils été responsables de tout ou partie des accumulations et de certaines modifications des faunes fossiles pléistocènes ? Ont-ils été en compétition avec les Hommes pour les proies et les cavités ? Correspondent-ils à des proies d’autres Carnivores ? Ont-ils pu faire partie des espèces exploitées par les Hommes au Paléolithique ? Ont-ils été domestiqués au Néolithique ? Cet article propose une synthèse de l’état des connaissances actuelles permettant de discuter de la place des Canidés dans les assemblages fauniques d’Afrique du Nord, en particulier au Pléistocène.
Matériel et méthodes
Ce travail s’appuie à la fois sur des données issues d’analyses taphonomiques récentes et sur des données provenant de références bibliographiques concernant les sites à Canidés à l’échelle de l’Afrique du Nord, depuis le Pléistocène jusqu’à l’Holocène. Concernant le rôle de ces Carnivores dans les accumulations de faune fossile, les études taphonomiques et archéozoologique restent rares et de qualité variée pour cette zone géographique. Pour les sites dont on dispose de données disponibles relativement complètes, les différents critères retenus pour la discussion sont : 1) les données générales : la localisation, les couches, la chronologie ; 2) le spectre faunique : les taux de Carnivores, de Canidés et d’Hyénidés, ainsi que des Gazelles; 3) les représentations anatomiques et les profils d’âge pour les Canidés et les Gazelles; 4) les données taphonomiques (traces de dents, restes semi-digérés, stries de boucherie, etc.…). Cette synthèse correspond à un état de l’art préliminaire et provisoire des connaissances actuelles concernant les sites à Canidés, amené à être précisé et complété dans les années à venir.
Présentation des données sur les sites à Canidés
Grotte à Hominidés (Pléistocène moyen)
La Grotte à Hominidés (GH) est localisée dans la région de Casablanca (Maroc) (Fig. 1). Comme son nom l’indique elle a livré des restes d’Hominidés, ainsi que des industries acheuléennes et une riche faune mammalienne datant d’environ 500 ka34. La liste faunique est diversifiée, comprenant des Bovidés, avec la prédominance des Antilopinés (Gazella cf. atlantica) et des Alcélaphinés (cf. Damaliscus)35. Les Carnivores sont abondants. Parmi ces derniers, les Canidés sont dominants et représentés principalement par un Chacal (Lupullela mohibi ; Fig. 3). Les Hyénidés sont présents, mais beaucoup plus rares et sont représentés par les deux taxons Crocuta crocuta et Hyaena hyaena. Parmi les Carnivores, nous remarquons aussi la présence de Panthera sp., Lynx sp. et Mellivora capensis. Les Gazelles et Alcélaphinés comptent des individus de tous âges, avec une prédominance des juvéniles. Les traces de dents de carnivores concernent environ 20% des restes au total (Fig. 3). Les dimensions des pits mettent en évidence l’intervention de Carnivores de tailles variées, avec une majorité de marques de petites dimensions attribuables au Chacal36. La taille des restes semi-digérés et des nombreux coprolithes recensés permet également d’identifier différentes catégories d’accumulateurs (Porc-épic, Chacal et Hyénidés). Aucune trace de découpe n’a été relevée. Un fragment diaphysaire de fémur humain porte des traces de dents de carnivore attribuées d’après leur localisation et leurs dimensions à un grand prédateur de type Hyénidé37. Cet assemblage faunique a été considéré comme le résultat d’occupations mixtes de Carnivores comprenant chacals, hyènes, grands et moyens Félidés et ours. Selon C. Daujeard et al.38, les petits Canidés pourraient avoir joué un rôle plus important que supposé antérieurement39 dans l’accumulation des restes osseux. R. Bernoussi s’interrogeait toutefois déjà sur le rôle de ce carnivore dans l’accumulation.
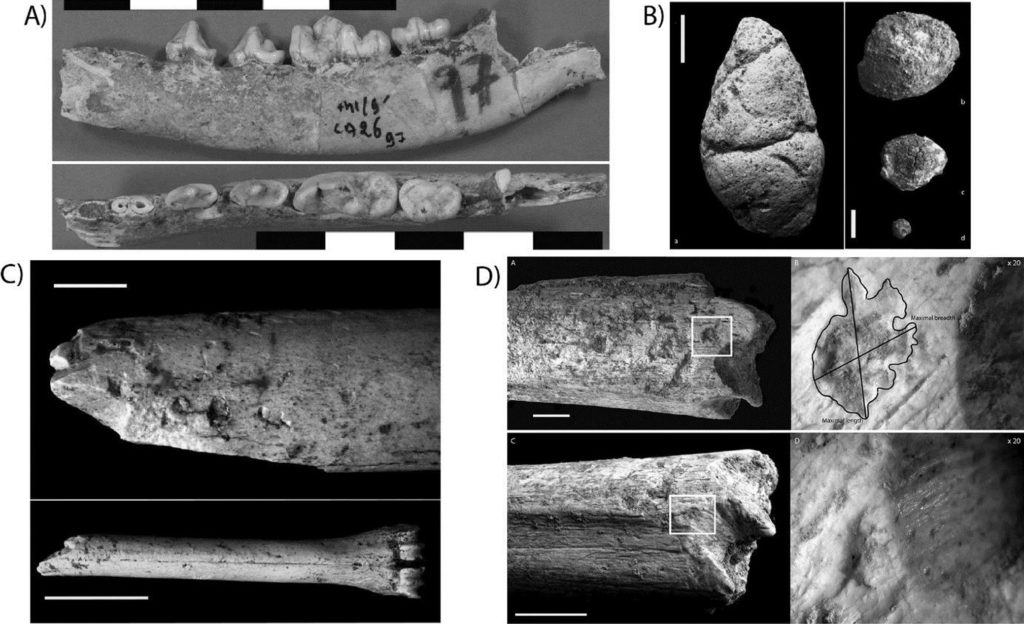
a. Hémi-mandibule de Lupulella mohibi (Cliché D. Geraads),
b. Différentes tailles de coprolithes, échelle : 1 cm (Daujeard et al. 2012),
c. Métacarpe d’Alcélaphiné avec des traces de mâchonnement, échelle : 1 cm (Daujeard et al. 2012),
d. Traces de dents de carnivore sur le fémur d’Hominidé, échelle : 1 cm (Daujeard et al. 2016).
Grotte des Rhinocéros (Pléistocène moyen)
La Grotte des Rhinocéros (GDR) est localisée également dans la région de Casablanca (Fig. 1). Elle a livré de nombreux restes de faune, notamment de Rhinocéros, associés à des industries acheuléennes40. Là aussi la faune est diversifiée, et très similaire à celle de GH. Les Carnivores sont assez nombreux, et c’est également Lupullela mohibi qui domine41. Parmi les Ongulés, les Gazelles et les Alcélaphinés sont majoritaires, aux côtés des Rhinocéros, dont l’abondance relative est la particularité de ce site. Comme à GH, les Gazelles et les Alcélaphinés sont représentés par toutes les classes d’âge et toutes les parties anatomiques. Les Rhinocéros comptent surtout des juvéniles et des individus âgés, et sont dominés par les restes crâniens. Les traces de Carnivores sont présentes pour environ 10% des restes au total et quelques stries de boucherie ont été mises en évidence42. Ainsi ce site correspondrait à un mélange d’occupations par les Hommes et les Carnivores, avec là aussi la question de l’intervention du chacal, charognard grignotant les restes abandonnés par les Hommes et/ou accumulateurs de certaines faunes (Gazelles par exemple) en parallèle à d’autres Carnivores dont les Hyénidés. Mais à l’inverse de la Grotte à Hominidés, la Grotte des Rhinocéros ne livre que très peu de coprolithes, de Carnivores juvéniles et de restes semi-digérés. La Grotte des Rhinocéros n’a donc certainement pas autant fonctionné en tanière que la Grotte à Hominidés.
Grotte d’El Harhoura 2 (Pléistocène supérieur)
La Grotte d’El Harhoura 2 est localisée à Témara, au Sud de Rabat (Maroc). La stratigraphie du porche de grotte comprend des niveaux du Pléistocène supérieur et de l’Holocène avec des industries attribuées à du Middle Stone Age, de l’Ibéromaurusien et du Néolithique43. La faune des niveaux de la fin du Pléistocène supérieur (couche 4A, 3 et 2) a livré un spectre diversifié largement dominé par les Antilopinés44. Selon les niveaux, les pourcentages de Carnivores oscillent entre 10 et 16%. Parmi les Carnivores, petits (Renard et/ou Chacal) et grands Canidés sont largement dominants (Fig. 4). L’Hyène est représentée par Crocuta crocuta45, qui représente seulement 0 à 1 % des Carnivores. Panthera sp. et Mellivora capensis sont aussi mentionnés. Les Gazelles comptent un nombre important de juvéniles et sont représentées par toutes les parties anatomiques. Le développement d’encroûtements sur les surfaces osseuses limite les observations taphonomiques. Cependant des traces de dents de Carnivore ont été observées sur 14 à 20% des restes au total (Fig. 4) et 6 à 9 % des restes sont semi-digérés. Ces derniers, tout comme les coprolithes, sont nombreux dans les refus de tamis. Ils sont de tailles variées, et appartiennent principalement à des Gazelles, mais aussi à de petits Canidés (Fig. 4). Ces restes semi-digérés sont probablement sous-estimés, car la fraction non-identifiée des refus de tamis n’a pas été considérée ici. Les restes osseux sont épars, même s’il y a quelques connexions. Les os longs sont fragmentés, principalement sur os frais, mais de nombreuses extrémités, en particulier pour les Gazelles, sont conservées. Notons la présence de rares traces de découpe (1-2%), de quelques restes brûlés présents en particulier dans la couche 4A où a été décrite une structure de combustion et de “retouchoirs”. Dans cette grotte, l’accumulation faunique est principalement attribuée à l’intervention des Carnivores, et en particulier aux Canidés, associée à quelques passages brefs des Hommes46.
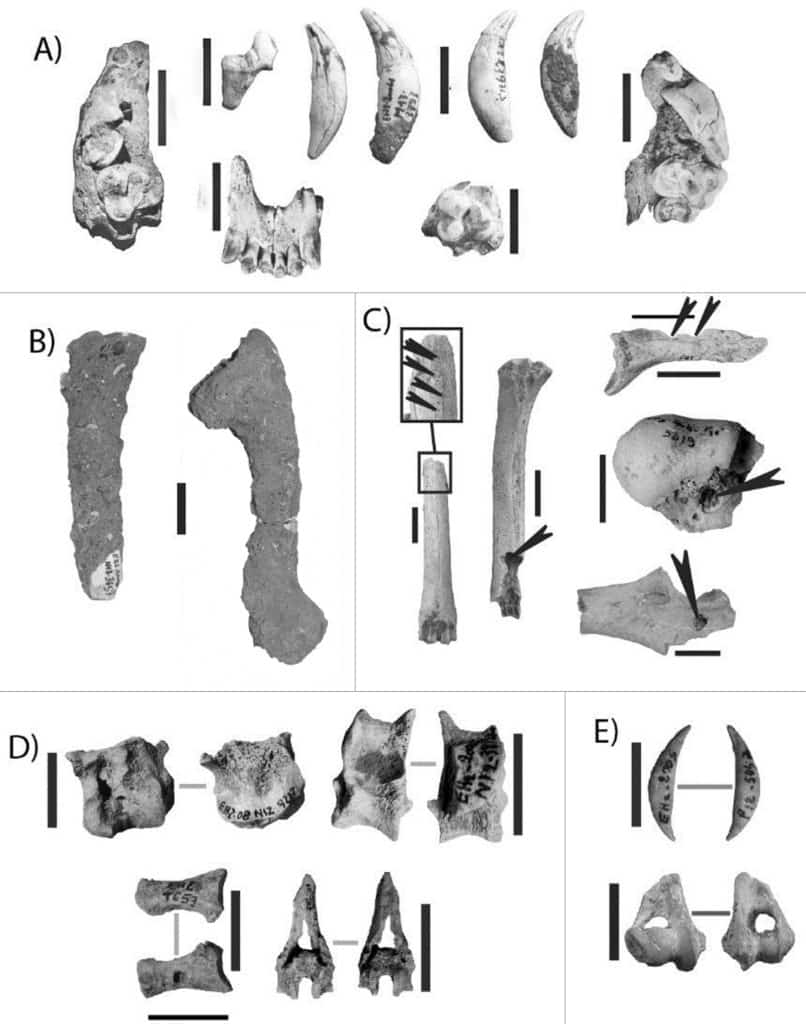
a. Restes de grand Canidé,
b. Restes de Gazelle encroûtés,
c. Restes de Gazelle avec des traces de dents,
d. Restes de Gazelle semi-digérés,
e. Restes de petit Canidé semi-digérés.
Échelles : 2 cm (Campmas et al. 2017).
Grotte des Gazelles (Pléistocène supérieur)
La grotte des Gazelles (Dar Bouazza, Tamaris) est située au Sud de Casablanca. Le remplissage de cette poche karstique est rapporté à la fin du Pléistocène. Elle a fourni des restes de faune accompagnés de quelques éléments lithiques attribués au Middle Stone Age (sans pièces pédonculées et qualifié d’Atéro-Moustérien) et à l’Ibéromaurusien47. Le remplissage est subdivisé en deux grands ensembles stratigraphiques. Le niveau supérieur est celui qui a livré la majorité du matériel faunique.
D’après Daujeard et al.48, la faune est largement dominée par les Gazelles appartenant à toutes les classes d’âge. Parmi les Carnivores, les Canidés représentent 90 % et comprennent deux taxons :
Vulpes vulpes et Canis “aureus” (cf. grand Canidé ; Fig. 5). La présence de l’Hyène, Hyaena hyaena, et de deux gros coprolithes, est documentée. Des restes en connexion anatomique sont décrits. Des traces de dents de Carnivore sont encore une fois présentes sur 17 % des restes et aucune trace de découpe n’a été relevée (Fig. 5). Bougariane et al.49 avaient obtenu des résultats sensiblement similaires, avec la présence de restes de Crocuta crocuta dans le locus principal. Concernant le grand Canidé, Bougariane et al. différencient l’espèce Canis aureus du grand Canis sp. et sur la base d’une étude paléontologique proposent d’attribuer ce dernier à un Chacal de grande stature ayant la taille du Loup50 (Fig. 5). Néanmoins, lors de la rédaction des articles de Bougariane et al., l’espèce Canis lupus lupaster/Canis anthus venait à peine d’être identifiée dans cette région du globe51, de sorte que ses dimensions et sa morphologie n’ont donc pas été intégrées à leurs comparaisons. Ce site correspondrait à un remplissage naturel d’aven-piège, associé à une intervention des Carnivores, principalement des Canidés, mais aussi de l’Hyène, avec peut-être des visites sporadiques des Hommes marquées par la présence des industries lithiques52.
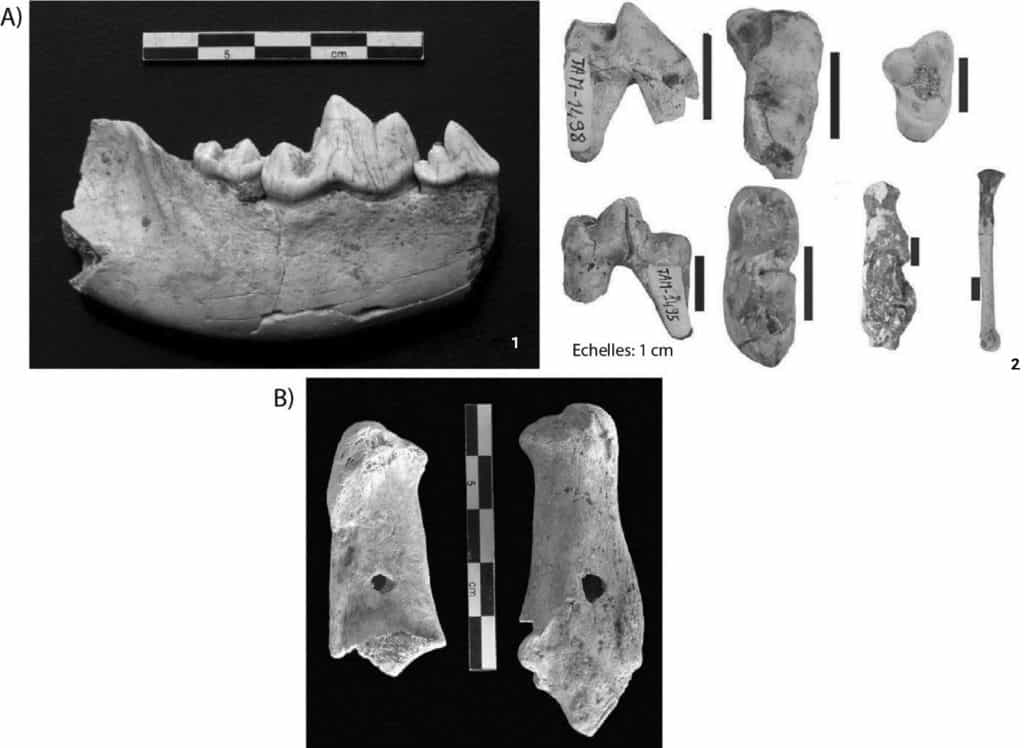
a. Restes de grand Canidé (1=(Daujeard et al. 2011), 2= (Bougariane et al. 2012)),
b. Traces de dents sur des calcanéums de Gazelle (Clichés C. Daujeard).
Grotte d’El Harhoura 1 (Pléistocène supérieur)
La grotte d’El Harhoura 1 est située à proximité d’El Harhoura 2 dans la région de Témara. Cette cavité a fait l’objet de fouilles de sauvetage en 1977 par A. Debénath. Associés à une faune pléistocène, des restes humains et des industries du Middle Stone Age (Atérien) y ont été récoltés. Comme dans les autres cavités, le spectre faunique est diversifié tout en étant dominé par les Bovidés53. Les pourcentages de Carnivores oscillent entre 12 et 20 %. Les Canidés, avec les taxons Canis “aureus” et Vulpes vulpes, sont bien représentés avec environ 70 % des Carnivores. Les Hyénidés, qui comptent de 1 à 7 % des Carnivores, sont représentées par Crocuta crocuta et Hyaena hyaena54. Parmi les Ongulés, les Gazelles dominent avec 80% des Ongulés55. Elles sont représentées là aussi par toutes les classes d’âge et toutes les portions anatomiques. Notons également la présence d’un grand nombre de Léporidés56. Les traces de dents de Carnivore ont été relevées sur 9 % des restes de Gazelle. Des restes semi-digérés, des coprolithes, ainsi que de très rares traces de découpe sont mentionnés. Cette accumulation faunique a été attribuée à une occupation mixte, principalement de Carnivores, en particulier d’Hyènes. L’Homme semble être passé dans la cavité (rares traces de découpe, industrie lithique et mollusques marins)57.
Grotte d’El Mnasra (Pléistocène supérieur)
La grotte d’El Mnasra est également localisée dans la région de Témara. Les fouilles se situent à l’intérieur de la cavité. Pour le Pléistocène, les restes de faunes discutés ici proviennent de la fouille de l’Unité Stratigraphique 4 (US 4), qui d’après l’étude géomorphologique, se serait déposée à la fin du Pléistocène, pendant une période d’aridification (MIS 4-2)58. L’étude préliminaire des faunes suggère que les Gazelles sont largement majoritaires avec plus de 90 % des Ongulés. De nombreux restes de Gazelle et de petits Canidés sont semi-digérés (23% des restes) (Fig. 6). Cette grande quantité de petits restes semi-digérés appartenant à des petites proies soulève donc là encore des questions sur la responsabilité des Canidés dans la formation de cet assemblage59.

a. De Gazelles,
b. De petits Canidés.
Échelles : 1 cm (Campmas et al. 2015).
Mugharet el Aliya (Pléistocène supérieur)
Le site de Mugharet el Aliya est situé dans la région de Tanger. Dans les niveaux du Middle Stone Age, le spectre est diversifié avec la présence de nombreux Canidés aux côtés des Hyénidés. Les faunes du niveau inférieur (L.9) sont attribuées à l’activité de l’Hyène, du fait de la fréquence des Carnivores (19 %), des nombreuses traces de mâchonnement (19 % des restes) et des nombreux restes d’Hyène (36 % des Carnivores) attribués à Crocuta crocuta, dont des juvéniles60. Toutefois, les Canidés, Canis “aureus” et Vulpes vulpes, sont encore mieux représentés (59 % des Carnivores), ce qui soulève ici des questions sur leur statut. Leur présence signe-t-elle une possible contribution de ces Canidés dans l’accumulation des restes de faune, ou ces Canidés sont-ils les proies d’autres prédateurs (Hyène) ? Dans les niveaux inférieurs 9 et 10, les Gazelles sont moins fréquentes que dans les autres sites (41 % des Ongulés), au profit des Équidés (42 % des Ongulés). On note là aussi la présence des Léporidés et de quelques Hystricidés61. Pour les niveaux supérieurs du Middle Stone Age (L.6-5), les Gazelles sont plus fréquemment identifiées (81 % des Ongulés). Dans ces niveaux, l’accumulation est plus largement attribuée à l’Homme du fait de la fréquence un peu plus élevée des traces de découpe (1-7 %), de la plus faible fréquence des Carnivores (11 %, dont 57 % de restes de Canidés) et du pourcentage légèrement plus faible de traces de dents que dans le niveau inférieur (4-7 %). Cependant, il est important de souligner ici que ces pourcentages sont assez similaires aux autres sites où l’intervention des Canidés semble attestée.
Grotte des Contrebandiers (Pléistocène supérieur)
D’après Aldeias et al. et Jacobs et al.62, la stratigraphie de la grotte des Contrebandiers présente une lacune sédimentaire entre les dépôts du stade isotopique 5 associés à du Middle Stone Age et l’Ibéromaurusien. Les restes de faune sont donc associés à des industries du Middle Stone Age (atériennes et moustériennes), ainsi qu’ibéromaurusiennes63.
L’étude taphonomique et archéozoologique de la faune a fait l’objet d’une thèse64. Le spectre faunique est diversifié avec 16 espèces d’Ongulés et 12 de Carnivores. La fréquence des Carnivores est très variable d’un niveau à l’autre. Dans tous les niveaux, les Canidés sont bien représentés (entre 30 et 80 % des Carnivores selon les niveaux), particulièrement par Vulpes rueppellii (42 % des Canidés), alors que ce taxon n’apparaît pas dans les listes fauniques des Carnivores de cette même région. Sont mentionnés aussi le Chacal, Canis “aureus”, une nouvelle espèce de Chacal, Canis nov. sp., le Renard des sables, Vulpes zerda, et le Lycaon, Lycaon pictus. Les Hyènes sont également présentes (jusqu’à 6 % des Carnivores selon les niveaux), surtout dans les niveaux atériens où l’activité humaine est bien enregistrée. Les Gazelles sont bien représentées, mais largement moins que dans les autres sites de la région (17-40 % des Ongulés). Toutefois les restes de mammifères et de Bovidés indéterminés de petite-moyenne taille sont nombreux.
Pour tous les niveaux, E. Yuko Hallett65 suggère que les Hommes sont à l’origine de l’accumulation des restes de faune et propose un accès secondaire des Carnivores. En effet, elle relève des taux de traces de découpe élevés, même parfois supérieurs à 50% sur les diaphyses des os longs. Des traces de découpe sont même observées sur des mandibules et des os longs de Carnivores, notamment dans les niveaux ibéromaurusiens. E. Yuko Hallett les attribue à la pratique de la pelleterie, notamment du fait de la faible fracturation des restes de Carnivore. Cependant, dans certains niveaux, notamment ceux du Paléolithique moyen, les pourcentages de traces de Carnivores sont plus élevés que ceux des marques anthropiques. De plus, la faible fracturation des restes de Carnivores ne peut-elle pas être induite par la difficulté d’identification des fragments leur appartenant, d’autant que les traces localisées sur les diaphyses des os longs peuvent faire penser aussi au retrait de la viande.
Tamar Hat (Phase finale du Pléistocène supérieur)
Les niveaux ibéromaurusiens de Tamar-Hat (Algérie) ont livré de la faune comprenant de nombreuses espèces. Contrairement aux sites de la côte atlantique, les faunes sont dominées par le Mouflon à manchettes (Ammotragus lervia). Les fréquences de Carnivores sont faibles (2 %) et ces derniers sont représentés seulement par les Canidés, le Chacal, Canis “aureus” et le Renard, Vulpes vulpes66. La présence d’une trace de découpe sur la partie diaphysaire d’une ulna de Renard est mentionnée et attribuée également à de la pelleterie67. Là encore, le retrait de la viande ne semble pas à exclure des possibilités d’interprétation de cette trace.
Discussion – Conclusion
Concernant la taxinomie des Canidés du Maghreb, selon les auteurs et les études les attributions spécifiques peuvent varier. Par exemple, les Canidés de la Grotte à Hominidés (Acheuléen) sont actuellement tous attribués à un Chacal de petite dimension Lupulella mohibi68. A l’Oued Sarrat, le grand Canidé associé à des industries acheuléennes est attribué à Canis othmanii sp. nov.69. Pour une partie des sites du Pléistocène supérieur de Témara au Maroc, comme pour les sites algériens de cette période, les grands Canidés sont identifiés comme appartenant à Canis aureus aux côtés du Renard70. Pour les plus grands spécimens de Canis aureus décrits à Témara, ils sont parfois attribués à un Canidé de grande dimension, Canis sp. ou Canis nov. sp.71.
Les données récentes, à la fois génétiques et morphologiques, suggèrent que le Chacal d’Afrique du Nord serait plus proche du Loup gris (Canis lupus) que du Chacal européen72. Il pourrait donc être attribué à Canis lupus lupaster73 ou à Canis anthus, le Chacal du Sénégal ou Loup africain74, plutôt qu’à Canis aureus. Les grands Canis fossiles seraient alors probablement leurs ancêtres75.
L’attribution taxinomique des petits Canidés soulève le même type de problématique. Ils rassemblent le Chacal africain du genre Lupulella dans les niveaux pléistocènes moyens à la Grotte à Hominidés et à la Grotte des Rhinocéros76. Au Pléistocène supérieur on trouve le Renard roux marocain (Vulpes vulpes atlantica) à Doukkala 277, le Renard roux (Vulpes vulpes) à El Harhoura 1, à la Grotte des Gazelles ou à Mugharet el Aliya78, les petits Canidés indéterminés – cf. Renard (Vulpes vulpes) / petit Chacal (Canis aureus et/ou Lupulella sp.) – à El Harhoura 2 et El Mnasra79 et le Renard de Rüppell (Vulpes rueppellii) et le Renard des sables (Vulpes zerda) aux Contrebandiers80. Actuellement, Vulpes rueppellii n’est décrit que dans le Sud du Maroc, tout comme Vulpes zerda. Le renard roux Vulpes vulpes est présent dans tout le Nord du Maroc81.
Concernant le statut des Canidés dans les assemblages de faunes fossiles d’Afrique du Nord, les données rassemblées ici permettent d’avoir quelques éléments de réponses :
Étaient-ils en compétition avec les Hommes ou d’autres Carnivores pour les proies et les cavités ?
C’est probablement le cas pour ce qui est du petit Chacal Lupulella présent au Pléistocène moyen à la Grotte à Hominidés et du grand Canidé dans les sites de Témara de la fin du Pléistocène supérieur. Cependant l’Hyène semble toujours avoir aussi joué un rôle dans la mise en place de ces assemblages fauniques82. Néanmoins, la bonne conservation des extrémités d’os longs dans les assemblages ne va pas dans le sens d’une intervention majoritaire des Hyénidés, au fort pouvoir destructeur. À l’exception des niveaux inférieurs de Mugharet el Aliya et du site de Tamar Hat, dans la plupart des sites pléistocènes du Maghreb qui comprennent de nombreux restes de Canidés, qu’il s’agisse de Lupulella mohibi ou de Canis aureus/Canis sp., les restes de petits Ongulés (Gazelles) sont très abondants. Les traces de dents sont présentes et les restes semi-digérés, lorsqu’ils sont décomptés, sont fréquents. La taille des proies ainsi que le nombre et la morphologie des restes semi-digérés rappellent ceux décrits dans les tanières de Cuons fossiles et de loups actuels83. En effet, les Canidés sont connus pour accumuler principalement des petites proies (micro- et mésofaune), et même des petits Herbivores tels que les Gazelles pour les plus grandes formes. À la liste proposée, nous pouvons probablement y rajouter, Doukkala 284, Dar es Soltane 185, les Grottes d’El Khenzira86), le Jebel Irhoud-niveaux supérieurs87 ou Khef-el-Baroud88, où les données moins précises mentionnent la présence de Canidés et d’Hyènes aux côtés d’un grand nombre de Gazelles, associée parfois à la mention de traces de dents de Carnivore, de restes semi-digérés ou de coprolithes. Il est à noter par ailleurs que le pourcentage plus important de grands ongulés (Équidés, grands Bovidés) dans des niveaux du Pléistocène supérieur de sites comme la Grotte des Félins89 ou Mugharet el Aliya, peut être due à la forte présence de grands Carnivores comme les Hyénidés ou les Félidés. Seuls de futurs travaux (poursuite des études taphonomiques et mise en place de référentiels actualistes) permettront de vérifier si les Canidés peuvent bien être comptés parmi les accumulateurs aux côtésde ces grands Carnivores et quelle est la part des accumulations liée à l’un ou à l’autre de ces prédateurs.
Correspondent-ils à des proies d’autres Carnivores ?
Si certains critères font des Canidés de potentiels accumulateurs, la présence de quelques-uns de leurs restes avec des traces de Carnivores (traces de dents et éléments semi-digérés) suggèrent qu’ils ont été aussi des proies. C’est le cas notamment du Renard à El Harhoura 290 ou du Chacal dans les sites des carrières Thomas91.
Étaient-ils utilisés (viande et/ou peau) par les Hommes ?
Les traces liées à une utilisation des Canidés par les Hommes sont rares au Pléistocène. La plupart du temps, ces marques sont associées à la pelleterie, avec notamment l’exemple des sites des Contrebandiers92 et de Tamar Hat93. Malgré tout, la localisation de certaines stries, par exemple sur la diaphyse d’un tibia de Vulpes rueppellii aux Contrebandiers ou sur la diaphyse d’une ulna de Vulpes vulpes à Tamar Hat, pourrait suggérer une consommation de la viande. À l’Holocène, plusieurs cas de Chiens portant des traces de consommation par l’Homme sont connus (communication personnelle de B. Bougariane).
Ont-ils été domestiqués ?
Les données sur la domestication des Canidés et la présence de restes de Chiens en Afrique du Nord-Ouest sont rares et la domestication n’est mentionnée qu’à partir de l’Holocène. B. Ouchaou & F. Amani94 fournissent une synthèse d’occupations holocènes (attribuées au Néolithique et à la Protohistoire) ayant livré des restes de Chien (Canis lupus familiaris) : Dar es Soltane 1, Grottes du cap Achakar, oued Merzeg, les Grottes du Rif Oriental, Kaf-That-el-Ghar, Kef el Baroud, Ghar-Khal, Bou-Saria, Abri Hassi Ouenzega. A ces sites se rajoutent El Harhoura 295, Guenfouda96 (Fig. 7), ainsi qu’en Algérie, une grotte près d’Alger, une autre près d’Oran et Djebel Roknia97. Les études paléontologiques suggèrent que ces Chiens étaient de taille moyenne à grande, en tout cas plus grande que le petit Chien européen. Ces Chiens sont en général présents aux côtés d’autres Carnivores sauvages, notamment des Canidés tels que le Renard et le Chacal98.
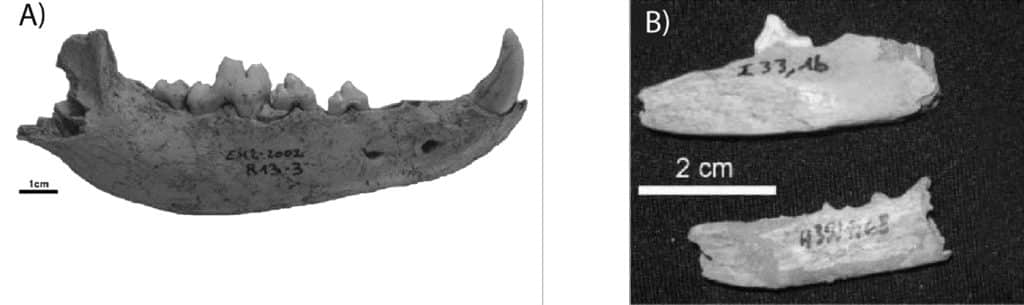
a. El Harhoura 2 (Stoetzel et al. 2012) ;
b. Guenfouda (Aouraghe et al. 2010).
Dans les niveaux holocènes de Guenfouda, probablement néolithiques, le Chien est accompagné d’autres faunes domestiques, la Chèvre, Capra hircus et le Mouton, Ovis aries99. La question de l’origine du Chien à l’Holocène, allochtone (Européenne ou Sub-Saharienne) ou autochtone (domestication in situ) n’est pas abordée. De la même façon, son rôle au sein des sociétés holocènes n’est pas précisé (aide à la chasse, gardien, nettoyeur de déchets, symbolique, ressource alimentaire, fournisseur de peau, animal de compagnie, animal défensif, etc.). Notons qu’à l’heure actuelle, les Canidés de cette période, notamment le Renard ou le Chien, n’ont été trouvés qu’en contexte sépulcral, associés à des squelettes humains comme dans des sites levantins ou chypriotes natoufiens e.g.100. Certains restes de Chien portent des traces de découpe ou ont été utilisés en élément de parure (communication personnelle de B. Bougariane). Certaines gravures de l’Atlas saharien renseignent également sur leur rôle, comme dans la représentation des “Chiens à la curée” de l’Oued Metlili (aide à la chasse ou nettoyeurs ?) (Fig. 8), ou aux côtés de représentations d’Équidés à Iche101 (gardien de troupeaux ?) ou de figurations humaines (probablement une femme et un enfant) à Adrar Metgourine près de Tata102 (animal de compagnie ?). On trouve également des figurations zoomorphes d’”Hommes-Chiens” en Lybie (symboliques ?)103 et à Volubilis une statuette (défense ?)104 et des mosaïques romaines avec des Chiens, dont un tenu en laisse (animal de compagnie ?)105. En Afrique du Nord, dans l’actuel, trois races de chien pourraient avoir des origines anciennes “Le lévrier de l’Azawakh”, le “Berger de l’Atlas, Chien Kabyle ou Chien des douars”, qui a une morphologie lupoide, et le “Chien de dénomination berbère”, qui est présent dans le langage berbère ancien106.
Pour conclure, les données taphonomiques et archéozoologiques peuvent parfois mettre en évidence les rôles multiples des Canidés au sein d’un même assemblage ou dans des sites subcontemporains différents. Ainsi, à Doukkala 2 ou El Harhoura 1, les Canidés sont plutôt identifiés comme les proies des Hyènes107. À la Grotte à Hominidés, El Harhoura 2 ou El Mnasra108, ils sont considérés comme des prédateurs responsables de l’accumulation des restes de Gazelles109. À la Grotte des Gazelles, les Canidés auraient visité la cavité pour consommer les carcasses tombées naturellement dans un aven110. Enfin, aux Contrebandiers, certains des Canidés présents seraient les proies des Hommes111.
Sans être exhaustive, cette synthèse illustre donc le fait que, tout comme en Europe, les Canidés ont probablement joué un rôle plus important qu’initialement proposé dans les accumulations et les modifications des faunes fossiles pléistocènes du Maghreb, en plus de celui des Hyénidés. Toutefois, les pistes de recherches identifiées ici doivent être maintenant approfondies par des analyses plus poussées et la multiplication des référentiels. Un réexamen poussé de ces assemblages de faunes fossiles, notamment pour les Canidés et les petits Bovidés, associé à la mise en place de référentiels actualistes sur les modalités de consommation des proies/charognes par les différents types de Canidés en comparaison aux autres Carnivores, est nécessaire (séquence de destruction, type de modifications, dimension des traces dents, fracturation, contenu coprogénique, degré de digestion, etc.). Même si des approches de ce type ont été entreprises depuis quelques temps en Europe112, elles demandent à être complétées et standardisées, en particulier pour les Canidés nord-africains113.
Enfin, seules des discussions et des confrontations devant le matériel permettront d’affiner et de valider les interprétations.

Remerciements
Cet article paraît un an après le décès d’Émilie Campmas, premier auteur, qui nous a quittés le 8 mars 2019. Nos remerciements vont à Myriam Boudadi-Maligne et Jean Baptiste Mallye (PACEA UMR 5199, France) pour leur invitation à participer au colloque “Relations Hommes-Canidés”, qui fait suite au projet “TeHoTeCa : Terre d’Homme, Terre de Canidés, évolution d’une interaction” financé par LaScArBx, et pour nous avoir permis cette publication. Nous remercions également les programmes de fouille de Casablanca et d’El Harhoura-Témara financés par le Ministère des Affaires Européennes et du Développement International (Paris) et le Ministère de la Culture (Maroc), ainsi que leurs directeurs, Jean Paul Raynal (PACEA UMR 5199, France), David Lefèvre (UMR 5140, France) et Abderrahim Mohib (Direction Régionale de la Culture, Maroc), ainsi que Mohamed Abdeljalil El Hajraoui (INSAP, Maroc) et Roland Nespoulet (HNHP UMR 7194, MNHN, France). Un grand merci à Bouchra Bougariane (Université de Meknès, Maroc) pour nous avoir indiqué que certains restes de Chiens holocènes portent des traces particulières, ainsi que Faysal Lemjidi (Université de Marrakech, Maroc) et Iddir Amara (CNRPAH, Algérie) pour la documentation sur les représentations rupestres. Nous remercions Denis Geraads pour sa relecture attentive, notamment en ce qui concerne les données sur la phylogénie des Canidés d’Afrique du Nord. Merci à Mathieu Lejay pour son aide dans la recherche et la compilation de fichiers appartenant à Émilie Campmas en lien avec cet article. Enfin, un grand merci aux deux rapporteurs, Sandrine Costamagno et Jean-Baptiste Mallye qui, grâce à leurs corrections et leurs commentaires pertinents, ont permis d’améliorer le manuscrit.
Bibliographie •••
- Aldeias, V., Goldberg, P., Dibble, H.L. et El Hajraoui, M.A. (2014) : “Deciphering site formation processes through soil micromorphology at Contrebandiers Cave, Morocco”, Journal of Archaeological Science, 69, 8-30.
- Amani, F. (1991) : La faune du gisement à Hominidés du Jebel Irhoud. Contribution à l’étude de la chronologie et de l’environnement du Quaternaire marocain, thèse de doctorat de l’Université Mohammed V, Rabat.
- Amri, L., Bartolini Lucenti, S., Mtimet, M.S., Karoui-Yaakoub, N., Ros-Montoya, S., Espigares, M.-P., Boughdiri, M., Bel Haj Ali, N. et Martínez-Navarro, B. (2017) : “Canis othmanii sp. nov. (Carnivora, Canidae) from the early Middle Pleistocene site of Wadi Sarrat (Tunisia)”, Comptes Rendus Palevol, 16, 774-782.
- Aouraghe, H. (2000) : “Les carnivores fossiles d’El Harhoura 1, Temara, Maroc”, L’Anthropologie, 104, 147-171.
- Aouraghe, H. (2001) : Contribution à la connaissance des faunes pléistocènes supérieures du Maroc: les vertébrés d’El Harhoura (Témara) comparés à ceux de plusieurs sites du Maghreb, thèse de doctorat de l’Université Mohamed I, Oujda.
- Aouraghe, H. (2004) : “Les populations de mammifères atériens d’El Harhoura 1 (Témara, Maroc)”, Bulletin d’Archéologie Marocaine, 20, 82-104.
- Aouraghe, H., Agusti, J., Ouchaou, B., Bailon, S., Lopez-Garcia, J.M., Haddoumi, H., Hammouti, K.E., Oujaa, A. et Bougariane, B. (2010) : “The Holocene vertebrate fauna from Guenfouda site, Eastern Morocco”, Historical Biology, 22, 320-326.
- Aouraghe, H., Bougariane, B. et Abbassi, M. (2012) : “Les lagomorphes du Pléistocène supérieur de la grotte d’El Harhoura 1 (Témara, Maroc) ”, Quaternaire, 23, 2, 163-174.
- Arilla, M., Rosell, J. et Blasco, R. (2019): “Contributing to characterise wild predator behaviour: consumption pattern, spatial distribution and bone damage on ungulate carcasses consumed by red fox (Vulpes vulpes)”, Archaeological and Anthropological Sciences, 11, 2271-2291.
- Aulagnier, S. (2008) : Guide des mammifères d’Europe, d’Afrique du Nord et du Moyen-Orient, Paris.
- Aulagnier, S., Cuzin, F. et Thévenot, M. (2017) : “Mammifères sauvages du Maroc”, Société Française pour l’Étude et la protection des Mammifères, Paris.
- Barton, R.N.E., Bouzouggar, A., Collcutt, S.N., Gale, R., Higham, T.F.G., Humphrey, L.T., Parfitt, S., Rhodes, E., Stringer, C.B. et Malek, F. (2005) : “The Late Upper Palaeolithic Occupation of the Moroccan Northwest Maghreb During the Last Glacial Maximum”, African Archaeological Review, 22, 77-100.
- Barton, R.N.E., Bouzouggar, A., Collcutt, S.N., Schwenninger, J.-L. et Clark-Balzan, L. (2009) : “OSL dating of the Aterian levels at Dar es-Soltan I (Rabat, Morocco) and implications for the dispersal of modern Homo sapiens”, Quaternary Science Reviews, 28, 1914-1931.
- Barton, R.N.E., Bouzouggar, A., Hogue, J.T., Lee, S., Collcutt, S.N. et Ditchfield, P. (2013) : “Origins of the Iberomaurusian in NW Africa: New AMS radiocarbon dating of the Middle and Later Stone Age deposits at Taforalt Cave, Morocco”, Journal of Human Evolution, 65, 266-281.
- Barton, R.N.E., Bouzouggar, A., Collcutt, S.N., Carrión Marco, Y., Clark-Balzan, L., Debenham, N.C. et Morales, J. (2016) : “Reconsidering the MSA to LSA transition at Taforalt Cave (Morocco) in the light of new multi-proxy dating evidence”, Quaternary International, 413, A, 36-49.
- Benecke, N. (1987) : “Studies on early dog remains from Northern Europe”, Journal of Archaeological Science, 14, 31-49.
- Bernoussi, R. (1997) : Contribution à l’étude paléontologique et observation archéozoologiques pour deux sites du Pléistocène moyen du Maroc Atlantique: la Grotte à Hominidés de la Carrière Thomas 1 et la Grotte des Rhinocéros de la Varrière Oulad Hamida 1 (Casablanca, Maroc), thèse de doctorat de l’Université Bordeaux 1.
- Boube-Piccot, C. (1969) : “Les bronzes antiques du Maroc. I”, in : La statuaire, ETAM IV, Rabat, (2 vol.).
- Boudadi-Maligne, M. (2010) : Les Canis pléistocènes du sud de la France : approche biosystématique, évolutive et biochronologique, thèse de doctorat, université de Bordeaux 1.
- Boudadi-Maligne, M., Mallye, J.-B., Langlais, M. et Barshay-Szmidt, C. (2012) : “Des restes de chiens magdaléniens à l’abri du Morin (Gironde, France). Implications socio-économiques d’une innovation zootechnique”, Paléo, 39-54.
- Boudadi-Maligne, Escarguel, G., Tresset, A. et Vigne, J.-D. (2018a) : “Du loup au chien, un point sur la question de la domestication au cours du Paléolithique”, Paléo, 271-279.
- Boudadi-Maligne, M., Mallye, J.-B., Castel, J.-C., Ferrié, J.-G., Crépin, L., Kuntz, D., Vercoutère, C., Soulier, M.-C. et Costamagno, S. (2018b) : “Loups, chiens et sociétés du Paléolithique supérieur”, in : Animal Symbolisé, Animal Exploité : Du Paléolithique à La Protohistoire, Actes Des Congrès Des Sociétés Historiques et Scientifiques, 11, Paris, 199-213.
- Bougariane, B., Zouhri, S., Ouchaou, B. et Oujaa, A. (2012) : “Description et position systématique du grand canidé de Tamaris I (Casablanca, Maroc)”, Quaternaire, 23, 2, 149-156.
- Bougariane, B., Zouhri, S., Ouchaou, B., Oujaa, A. et Boudad, L. (2010): “Large mammals from the Upper Pleistocene at Tamaris I ‘Grotte des Gazelles’ (Casablanca, Morocco): paleoecological and biochronological implications”, Historical Biology, 22, 295-302.
- Bouzouggar, A. et Barton, N. (2012): “The identity and timing of the Aterian in Morocco”, in : Modern Origins, 93-105.
- Bouzouggar, A., Barton, R.N.E., Blockley, S., Bronk-Ramsey, C., Collcutt, S.N., Gale, R., Higham, T.F.G., Humphrey, L.T., Parfitt, S., Turner, E. et Ward, S. (2008) : “Reevaluating the Age of the Iberomaurusian in Morocco”, African Archaeological Review, 25, 3-19.
- Bouzouggar, A., Barton, N., Vanhaeren, M., d’Errico, F., Collcutt, S., Higham, T., Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J.-L., Stringer, C., Turner, E., Ward, S., Moutmir, A. et Stambouli, A. (2007) : “82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior”, PNAS, 104, 9964-9969.
- Bouzouggar, A., Humphrey, L., Barton, N., Parfitt, S., Clark-Balzan, L., Schwenninger, J.L., El Hajraoui, M.A., Nespoulet, R. et Bello, S.M. (2018) : “90,000 year-old specialised bone technology in the Aterian Middle Stone Age of North Africa”, PlosOne, 13, e0202021.
- Brugal, J.-P. et Boudadi-Maligne, M. (2011) : “Quaternary small to large canids in Europe: Taxonomic status and biochronological contribution”, Quaternary International, 243, 171-182.
- Campmas, É. (2012) : Caractérisation de l’occupation des sites de la région de Témara (Maroc) au Pléistocène supérieur et nouvelles données sur la subsistance des hommes du Paléolithique moyen d’Afrique du Nord: Exemples des approches taphonomiques et archéozoologiques menées sur les faunes d’El Harhoura 2 et d’El Mnasra, thèse de doctorat de l’Université de Bordeaux 1.
- Campmas, É. (2017) : “Integrating Human-Animal Relationships into New Data on Aterian Complexity: a Paradigm Shift for the North African Middle Stone Age”, African Archaeological Review, 34, 469-491.
- Campmas, É. et Beauval, C. (2008) : “Consommation osseuse des carnivores : résultats de l’étude de l’exploitation de carcasses de bœufs (Bos taurus) par des loups captifs”, Annales de Paléontologie, 94, 167-186.
- Campmas, É., Michel, P., Costamagno, S., Amani, F., Stoetzel, E., Nespoulet, R. et El Hajraoui, M.A. (2015): “Were Upper Pleistocene human/non-human predator occupations at the Témara caves (El Harhoura 2 and El Mnasra, Morocco) influenced by climate change? ”, Journal of Human Evolution, 78, 122-143.
- Campmas, É., Michel, P., Costamagno, S., El Hajraoui, M.A. et Nespoulet, R. (2017) : “Which predators are responsible for faunal accumulations at the Late Pleistocene layers of El Harhoura 2 Cave (Témara, Morocco)?”, Comptes Rendus Palevol, 16, 333-350.
- Campmas, É., Stoetzel, E. et Denys, C. (2018) : “African carnivores as taphonomic agents: Contribution of modern coprogenic sample analysis to their identification”, International Journal of Osteoarchaeology, 28, 237-263.
- Castel, J.C. (2004) : “L’Influence des canidés sur la formation des ensembles archéologiques : caractérisation des destructions dues aux loups”, Revue de Paléobiologie, 23, 675-693.
- Célérier, G., Tisnerat, N. et Valladas, H. (1999) : “Données nouvelles sur l’âge des vestiges de chien à Pont d’Ambon, Bourdeilles (Dordogne)/New data on the age of Canis remains at Pont d’Ambon, Bourdeilles (Dordogne, France)”, Paléo, 11, 163-165.
- Chaix, L. (2000) : “A preboreal dog from the Northern Alps (Savoie, France)”, in : Dogs through time: an archaeological perspective, Victoria-1998, BAR Int. Ser. 889, 49-59.
- Dachy, T., Guéret, C., Campmas, É., Simonnet, R. et Perrin, T. (2018) : “Saint-Trivier/Chabet el Houidga (Mascara, Algérie) : nouvel éclairage sur un faciès méconnu du début de l’Holocène”, Bulletin de la Société Préhistorique Française, 115, 215-251.
- Daujeard, C., Geraads, D., Raynal, J.-P., Mohib, A. et Gallotti, R. (2011) : “Carnivores et/ou hommes dans deux sites moustéro-atériens de Dar Bouazza (Casablanca, Maroc) : les données de la taphonomie”, in : Prédateurs Dans Tous Leurs États : Évolution, Biodiversité, Interactions, Mythes, Symboles, Actes des Rencontres Internationales d’Archéologie et d’Histoire d’Antibes, 2010, Antibes, 351-366.
- Daujeard, C., Geraads, D., Gallotti, R. and Raynal, J.-P. (2012) : “Carcass Acquisition and Consumption by Carnivores and Hominins in Middle Pleistocene Sites of Casablanca (Morocco)”, Journal of Taphonomy, 10, 349-372.
- Daujeard, C., Geraads, D., Gallotti, R., Lefèvre, D., Mohib, A., Raynal, J.-P. and Hublin, J.-J. (2016) : “Pleistocene Hominins as a Resource for Carnivores: A c. 500,000-Year-Old Human Femur Bearing Tooth-Marks in North Africa (Thomas Quarry I, Morocco)”, PlosOne, 11, e0152284.
- Daujeard, C., Falguères, C., Shao, Q., Geraads, D., Hublin, J. J., Lefevre, D., El Graoui, M., Rué, M., Gallotti, R., Delvigne, V., Queffelec, A., Ben Arous, E., Tombret, O., Mohib, A. et Raynal, J.-P. (2020) : « Earliest African evidence of carcass processing and consumption in cave at 700 ka, Casablanca, Morocco », Scientific reports, 10(1), 1-15.
- Dayan, T. (1994) : “Early Domesticated Dogs of the Near East”, Journal of Archaeological Science, 21, 633–640.
- Dibble, H., Aldeias, V., Alvarez-Fernandez, E., Blackwell, B., Hallett-Desguez, E., Jacobs, Z., Goldberg, P., Lin, S., Morala, A., Meyer, M., Olszewski, D., Reed, K., Rezek, Z., Richter, D., Roberts, R., Sandgathe, D., Schurmans, U., Skinner, A., Steele, T. et El-Hajraoui, M. (2012) : “New excavations at the site of Contrebandiers Cave, Morocco”, PaleoAnthropology, 145–201.
- Dibble, H.L., Aldeias, V., Jacobs, Z., Olszewski, D.I., Rezek, Z., Lin, S.C., Alvarez-Fernández, E., Barshay-Szmidt, C.C., Hallett-Desguez, E., Reed, D., Reed, K., Richter, D., Steele, T.E., Skinner, A., Blackwell, B., Doronicheva, E. et El-Hajraoui, M. (2013) : “On the industrial attributions of the Aterian and Mousterian of the Maghreb”, Journal of Human Evolution, 64, 194–210.
- El Hajraoui, M.A., Nespoulet, R., Debénath, A. et Dibble, H.L. (2012) : “Préhistoire de la région de Rabat-Témara”, Villes et Sites Archéologiques du Maroc (VESAM), Institut National des Sciences de l’Archéologie et du Patrimoine, Rabat, III.
- d’Errico, F., Vanhaeren, M., Barton, N., Bouzouggar, A., Mienis, H., Richter, D., Hublin, J.-J., McPherron, S. et Lozouet, P. (2009) : “Additional evidence on the use of personal ornaments in the Middle Paleolithic of North Africa”, PNAS, 106, 16051-16056.
- Espérandieu, G., Coppé, E.B. et Chaker, S. (1994) : “Chien”, in : Gabriel Camps, éd., Chèvre – Columnatien, Aix-en-Provence, Edisud, vol. 13.
- Ferguson, W.W. (1981) : “The systematic position of Canis aureus lupaster (Carnivora : Canidae) and the occurrence of Canis lupus in North Africa, Egypt and Sinai”, Mammalia, 45, 459-466
- Flower, L.O.H. and Schreve, D.C. (2014) : “An investigation of palaeodietary variability in European Pleistocene canids”, Quaternary Science Reviews, 96, 188-203.
- Fosse, P., Parrenin, F., Selva, N., Wajrak, A. et (2004) : “Premières observations néotaphonomiques sur des assemblages osseux de Bialowieza (nord-est Pologne) ”, Paléo, 16, 91-115.
- Fosse, P., Wajrak, A., Fourvel, J.B., Madelaine, S., Esteban-Nadal, M., Cáceres, I., Yravedra, J., Brugal, J.P., Prucca, A. et Haynes, G. (2012): “Bone Modification by Modern Wolf (Canis lupus): A Taphonomic Study From their Natural Feeding Places”, Journal of Taphonomy, 10, 197-217.
- Fourvel, J.B., Magniez, P., Moigne, A.-M., Testu, A., Joris, A., Lamglait, B., Vaccaro, C. et Fosse, P. (2018) : “Wild dogs and their relatives: implication of experimental feedings in their taphonomical identification”, Paléo, 29, 21-29.
- Gaubert, P., Bloch, C., Benyacoub, S., Abdelhamid, A., Pagani, P., Djagoun, C.A.M.S., Couloux, A. et Dufour, S. (2012) : “Reviving the African Wolf Canis lupus lupaster in North and West Africa: A Mitochondrial Lineage Ranging More than 6,000 km Wide”, PLoS One, 7, e42740.
- Geraads, D. (2002) : “Plio-Pleistocene Mammalian Biostratigraphy of Atlantic Morocco”, Quaternaire, 13, 43-53.
- Geraads, D. Raynal, J.-P. et Eisenmann V. (2002) : “The earliest human occupation of North Africa: a reply to Sahnouni et al. (2002)”, Journal of Human Evolution, 46, 751-761.
- Geraads, D. (2011) : “A revision of the fossil Canidae (Mammalia) of north-western Africa”, Palaeontology, 54, 429-446.
- Germonpré, M., Sablin, M.V., Stevens, R.E., Hedges, R.E.M., Hofreiter, M., Stiller, M. et Després, V.R. (2009) : “Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes”, Journal of Archaeological Science, 36, 473-490.
- Germonpré, M., Lázničková-Galetová, M. et Sablin, M.V. (2012) : “Palaeolithic dog skulls at the Gravettian Předmostí site, the Czech Republic”, Journal of Archaeological Science, 39, 184-202.
- Horard-Herbin, M.-P., Tresset, A. et Vigne, J.-D. (2014): “Domestication and uses of the dog in western Europe from the Paleolithic to the Iron Age”. Animal Frontiers, 4, 3, 23-31.
- Hublin, J.-J. (2009) : “The origin of Neandertals”, PNAS, 106, 16022-16027.
- Hublin, J.-J., Tillier, A.M. et Texier, J.-P. (1987) : “L’humérus d’enfant moustérien (Homo 4) du Djebel Irhoud (Maroc) dans son contexte archéologique”. Bulletins et Mémoires de la Société d’Anthropologie, XIV, 115-141.
- Hublin, J.-J., Ben-Ncer, A., Bailey, S.E., Freidline, S.E., Neubauer, S., Skinner, M.M., Bergmann, I., Le Cabec, A., Benazzi, S., Harvati, K. et Gunz, P. (2017) : “New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens”, Nature, 546, 289-292.
- Humphrey, L.T., Bello, S.M., Turner, E., Bouzouggar, A. et Barton, N. (2012) : “Iberomaurusian funerary behaviour: Evidence from Grotte des Pigeons, Taforalt, Morocco”, Journal of Human Evolution, 62, 261-273.
- Jacobs, Z., Meyer, M.C., Roberts, R.G., Aldeias, V., Dibble, H. et El Hajraoui, M.A. (2011) : “Single-grain OSL dating at La Grotte des Contrebandiers (‘Smugglers’ Cave’), Morocco: improved age constraints for the Middle Paleolithic levels”, Journal of Archaeological Science, 38, 3631-3643.
- Jacobs, Z., Roberts, R.G., Nespoulet, R., El Hajraoui, M.A. et Debénath, A. (2012) : “Single-grain OSL chronologies for Middle Palaeolithic deposits at El Mnasra and El Harhoura 2, Morocco: Implications for Late Pleistocene human–environment interactions along the Atlantic coast of northwest Africa”, Journal of Human Evolution, 62, 377-394.
- Koepfli, K.-P., Pollinger, J., Godinho, R., Robinson, J., Lea, A., Hendricks, S., Schweizer, R.M., Thalmann, O., Silva, P., Fan, Z., Yurchenko, A.A., Dobrynin, P., Makunin, A., Cahill, J.A., Shapiro, B., Álvares, F., Brito, J.C., Geffen, E., Leonard, J.A., Helgen, K.M., Johnson, W.E., O’Brien, S.J., Van Valkenburgh, B. et Wayne, R.K. (2015) : “Genome-wide Evidence Reveals that African and Eurasian Golden Jackals Are Distinct Species”, Current Biology, 25, 2158-2165.
- Lemjidi, F. (2018) : Contribution à l’étude des gravures rupestres de l’Anti-Atlas Occidental, analyse, interprétation, Chronologie et Mise en Patrimoine, thèse de doctorat de l’Université de Cadi Ayyad, Marrakech.
- Lenoble, A. (2010) : “Lithostratigraphie de la grotte d’El Mnasra. Description et hypothèses proposées”, in: Mission Archéologique El Harhoura-Témara, El Hajraoui, 14-15.
- Lenoble, A. (2011) : “Étude géologique de la grotte d’El Mnasra”, Mission Archéologique El Harhoura-Témara, El Hajraoui, 8-11.
- Limane, H., Rebuffat, R. et Drocourt, D. (1998) : Volubilis : De mosaïque à mosaïque, Casablanca.
- Linstädter, J. (2008): “The Epipalaeolithic-Neolithic-Transition in the Mediterranean region of Northwest Africa”, Quartär, 55, 33-54.
- Linstädter, J., Eiwanger, J., Mikdad, A. et Weniger, G.-C. (2012) : “Human occupation of Northwest Africa: A review of Middle Palaeolithic to Epipalaeolithic sites in Morocco”, Quaternary International, 274, 158-174.
- Maher, L.A., Stock, J.T., Finney, S., Heywood, J.J.N., Miracle, P.T. et Banning, E.B. (2011) : “A unique Human-Fox burial from a Pre-Natufian cemetery in the Levant (Jordan)”, PlosOne, 6, e15815.
- Mallye, J.-B., Cochard, D. et Laroulandie, V. (2008) : “Accumulations osseuses en périphérie de terriers de petits carnivores : les stigmates de prédation et de fréquentation”. Annales de Paléontologie, 94, 187-208.
- Mallye, J.-B., Costamagno, S., Boudadi-Maligne, M., Prucca, A., Laroulandie, V., Thiébault, C. et Mourre, V. (2012) : “Dhole (Cuon alpinus) as a Bone Accumulator and New Taphonomic Agent? The Case of Noisetier Cave (French Pyrenees)”, Journal of Taphonomy, 10, 317-347.
- Merzoug, S. (2005) : Comportement de subsistance des Ibéromaurusiens d’après l’analyse archéozoologique des mammifères de Tamar Hat, Taza 1 et Columata (Algérie), thèse de Doctorat du Muséum National d’histoire Naturelle, Paris.
- Merzoug, S. et Sari, L. (2008) : “Re-examination of the Zone I Material from Tamar Hat (Algeria): Zooarchaeological and Technofunctional Analyses”, African Archaeological Review, 25, 57-73.
- Michel, P. et Wengler, L. (1993a) : “Le site paléontologique et archéologique de Doukkala II (Maroc, Pléistocène moyen et supérieur) : premier jalon en Afrique du Nord d’un comportement humain assimilable à un charognage contrôlé et actif”, Comptes Rendus de l’Académie des Sciences, Série 2, 317, 4, 557-562.
- Michel, P. et Wengler, L. (1993b) : “Un site paléontologique avec des vestiges archéologiques : La carrière Doukkala II (Région de Temara, Maroc atlantique) – Paléoécologie des faunes et contribution à la connaissance du comportement humain”. Paléo, 5, 11-41.
- Michel, P., Campmas, É., Stoetzel, E., Nespoulet, R., Abdeljalil El Hajraoui, M. et Amani, F. (2009) : “La macrofaune du Pléistocène supérieur d’El Harhoura 2 (Témara, Maroc) : données préliminaires”, L’Anthropologie, 113, 283-312.
- Michel, P., Campmas, E., Stoetzel, E., Nespoulet, R., Hajraoui, M.A.E. et Amani, F. (2010) : “Upper Palaeolithic (layer 2) and Middle Palaeolithic (layer 3) large faunas from El Harhoura 2 Cave (Témara, Morocco): paleontological, paleoecological and paleoclimatic data”, Historical Biology, 22, 327-340.
- Monchot, H. et Aouraghe, H. (2009) : “Deciphering the taphonomic history of an Upper Paleolithic faunal assemblage from Zouhrah Cave/El Harhoura 1, Morocco”, Quaternaire, 20, 2, 239-253.
- Musil, R. (2000) : “Evidence for the domestication of wolves in central European Magdalenian sites”, in : Dogs through time: an archaeological perspective, Victoria-1998, BAR Int. Ser. 889, 21-28.
- Nami, M. et Moser, J. (2010) : “La grotte d’Ifri n’Ammar : Le Paléolithique moyen”, Forschungenzur Archäologie Außereuropäischer Kulturen, Verlag éd.
- Nespoulet, R., Debénath, A., El Hajraoui, M.A., Michel, P., Campmas, Emilie, Oujaa, A., Ben-Ncer, A., Lacombe, J.-P., Stoetzel, E. et Boudad, L. (2008) : “Le contexte archéologique des restes humains atériens de la région de Rabat-Témara (Maroc): Apport des fouilles des grottes d’El Mnasra et d’El Harhoura 2”, in : Le Quaternaire marocain dans son contexte méditerranéen, Actes des quatrièmes rencontres des Quaternaristes marocains (RQM4), Oujda, Novembre 2007, 356-375.
- Ouchaou, B. et Amani, F. (2002) : “Les carnivores des gisements néolithiques et protohistoriques du nord du Maroc”, Quaternaire, 13, 79-87.
- Ouchaou, B., Amani, F. et Mouchine, T. (1998) : “Etude archéozoologique du gisement de Kef-el-Baroud”, Préhistoire Anthropologie Méditerranéennes, 78, 2738.
- Pionnier-Capitan, M., Bemilli, C., Bodu, P., Célérier, G., Ferrié, J.-G., Fosse, P., Garcià, M. et Vigne, J.-D. (2011) : “New evidence for Upper Palaeolithic small domestic dogs in South-Western Europe”, Journal of Archaeological Science, 38, 9, 2123-2140.
- Rabhi, M. (2009) : “Étude de l’industrie lithique du niveau ‘A’ du site de Ain Hanech : Approche expérimentale”, Athar, Revue scientifique d’archéologie et du patrimoine, 8, 13-37.
- Raynal, J.-P. et Mohib, A., éd. (2016) : Préhistoire de Casablanca. I – La Grotte des Rhinocéros (fouilles 1991 et 1996), Villes et Sites Archéologiques du Maroc (VESAM), Institut National des Sciences de l’Archéologie et du Patrimoine, Rabat, VI.
- Raynal, J.-P., Geraads, D., Magoga, L., El Hajraoui, M.A., Texier, J-P., Lefèvre, D. et Sbihi-Alaoui, F.-Z. (1993) : “La grotte des Rhinocéros (Carrière Oulad Hamida 1, anciennement Thomas III, Casablanca), nouveau site acheuléen du Maroc Atlantique”, Comptes Rendus de l’Académie des Sciences, 316, 1477-1483.
- Raynal, J.-P., Sbihi Alaoui, F.Z., Geraads, D., Magoga, L. et Mohib, A. (2001) : “The earliest occupation of North-Africa: the Moroccan perspective”, Quaternary International, 75, 65-75.
- Richter, D., Moser, J., Nami, M., Eiwanger, J. et Mikdad, A. (2010) : “New chronometric data from Ifri n’Ammar (Morocco) and the chronostratigraphy of the Middle Palaeolithic in the Western Maghreb”, Journal of Human Evolution, 59, 672-679.
- Richter, D., Grün, R., Joannes-Boyau, R., Steele, T.E., Amani, F., Rué, M., Fernandes, P., Raynal, J.-P., Geraads, D., Ben-Ncer, A., Hublin, J.-J. et McPherron, S.P. (2017) : “The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age”, Nature, 546, 293-296.
- Rodrigue, A. (1993) : “Documents rupestres de l’Adrar Metgourine (Maroc saharien). s, 1993”, Société d’études et de recherches préhistorique, 42, 49-56.
- Rueness, E.K., Asmyhr, M.G., Sillero-Zubiri, C., Macdonald, D.W., Bekele, A., Atickem, A. et Stenseth, N.C. (2011) : “The Cryptic African Wolf: Canis aureus lupaster Is Not a Golden Jackal and Is Not Endemic to Egypt”. PLoS ONE 6, e16385.
- Ruhlmann, A. (1951) : La grotte Préhistorique de Dar es-Sotlan.
- Ruhlmann, A. (1936) : Les grottes préhistoriques d’”El Khenzira” (région de Mazagan) : Contribution à l’étude du Paléolithique marocain (moyen et supérieur), Nogent-le-Rotrou.
- Sahnouni, M. (2005) : “Point des connaissances du Paléolithique ancien d’Afrique du Nord et la question de la première occupation humaine au Maghreb”, in : Le Paléolithique en Afrique : L’Histoire la plus longue, Paris, 99-128.
- Sahnouni, M., de Heinzelin, J., van der Made, J., Brown, F., Everett, M., Harichane, Z., Kandi, N., Rossell, J., Hadjouis, D., Derradji, A., Ollé, A., María Vergès, J., Medig, M. et Canals, A. (2013) : “La séquence plio-pléistocène d’Ain Boucherit-Ain Hanech (Algérie orientale): Biochronologie, environnements, et comportements des Hominidés”, in : Actes Du Colloque International de l’Afrique, Berceau de l’Humanité: Découvertes Récentes (Sérif, 26-28 Octobre 2009), CNRPAH, Algérie, 189-210.
- Sahnouni, M., Parés, J.M., Duval, M., Cáceres, I., Harichane, Z., Made, J. van der, Pérez-González, A., Abdessadok, S., Kandi, N., Derradji, A., Medig, M., Boulaghraif, K. et Semaw, S. (2018) : “1.9-million- and 2.4-million-year-old artifacts and stone tool–cutmarked bones from Ain Boucherit, Algeria”. Science, 362, 6420, 1297-1301.
- Sari, L. (2014): “Technological change in Iberomaurusian culture: The case of Tamar Hat, Rassel and Columnata lithic assemblages (Algeria)”, Quaternary International, 320, 131-142.
- Sari, L. et Kim, K.-J. (2017) : “Lithic Economy and Specialized Activities Among the Iberomaurusian Populations of Tamar Hat Rockshelter (Northeastern Algeria)”, African Archaeological Review, 34, 543-556.
- Scerri, E.M.L. (2013) : “The Aterian and its place in the North African Middle Stone Age”, Quaternary International, 300, 111-130.
- Simoneau, A. (1977) : Catalogue des sites rupestres du Sud marocain, -Rabat.
- Stoetzel, E., Bougariane, B., Campmas, É., Ouchaou, B. et Michel, P. (2012) : “Faunes et Paléoenvironnements”, in: Préhistoire de La Région de Rabat-Témara, Villes et Sites Archéologiques du Maroc (VESAM), Institut National des Sciences de l’Archéologie et du Patrimoine, Rabat, III, 35-51.
- Tchernov, E. et Valla, F.F. (1997): “Two New Dogs, and Other Natufian Dogs, from the Southern Levant”, Journal of Archaeological Science, 24, 65-95.
- Urios, V., Donat-Torres, M.P., Ramírez, C., Monroy-Vilchis, O. et Rgribi-Idrissi, H. (2015) : “El análisis del genoma mitocondrial del cánido estudiado en Marruecos manifiesta que no es ni lobo (Canis lupus) ni chacal euroasiático (Canis aureus)”, AltoterO, 3: 1-24.
- Val, A. et Mallye, J.-B. (2011) : “Small Carnivore Skinning by professionals: Skeletal Modifications and Implications for the European Upper Palaeolithic”, Journal of Taphonomy, 9, 221-243.
- Valla, F.R. (1975) : “La Sépulture H. 104 de Mallaha (Eynan) et le problème de la domestication du chien en Palestine”, Paléorient, 3, 287-292.
- van Albada, A. et van Albada, A.-M. (2000) : La Montagne des hommes-chiens, Paris.
- van de Loosdrecht, M., Bouzouggar, A., Humphrey, L., Posth, C., Barton, N., Aximu-Petri, A., Nickel, B., Nagel, S., Talbi, E.H., Hajraoui, M.A.E., Amzazi, S., Hublin, J.-J., Pääbo, S., Schiffels, S., Meyer, M., Haak, W., Jeong, C. et Krause, J. (2018) : “Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations”, Science, 360, 6388, 548-552.
- Vigne, J.-D. (2005) : “L’humérus de chien magdalénien de Erralla (Gipuzkoa, Espagne) et la domestication tardiglaciaire du loup en Europe)”, MUNIBE, 57, 279-287.
- Vigne, J.-D. et Guilaine, J. (2004) : “Les premiers animaux de compagnie, 8500 ans avant notre ère ? … ou comment j’ai mangé mon chat, mon chien et mon renard”, Anthropozoologica, 39, 249-273.
- Viranta, S., Atickem, A., Werdelin, L. et Stenseth, N.C. (2017) : “Rediscovering a forgotten canid species”, BMC Zoology, 2, 6.
- Waters, S., El Harrad, H., Amhaouch, Z., Taiqui, L. et Senn, H. (2015) : “DNA analysis confirms African wolf in Morocco Canid”, Biology & Conservation, 18, 15-17.
- Wrinn, P.J. (2001) : “Reanalysis of the Pleistocene Archaeofauna from Mugharet El Aliya, Tangier, Morocco: implications for the Aterian”, in : UISPP Congress, Liège.
- Yuko Hallett, E. (2018) : Zooarchaeological and Taphonomic Analyses of Pleistocene Vertebrate Fauna from the Middle and Later Stone Age Occupations at Contrebandiers Cave, Atlantic Coast, Morocco, thèse de doctorat de l’Université d’Arizona State University.
Notes
- Germonpré et al. 2009 ; Boudadi-Maligne 2010 ; Brugal & Boudadi-Maligne 2011 ; Flower & Schreve 2014.
- Mallye et al. 2012.
- Mallye et al. 2008 ; Mallye ce volume ; Leoni et al. ce volume.
- Val & Mallye 2011.
- Benecke 1987 ; Dayan 1994 ; Tchernov & Valla 1997 ; Célérier 1999 ; Chaix 2000 ; Musil 2000 ; Vigne 2005 ; Pionnier-Capitan et al. 2011 ; Germonpré et al. 2012 ; Boudadi-Maligne et al. 2012, 2018a,b ; Horard-Herbin et al. 2014.
- Geraads 2011 ; Bougariane et al. 2012 ; Amri et al. 2017.
- Geraads 2011.
- Sahnouni et al. 2018.
- Geraads et al. 2002.
- Sahnouni et al. 2013.
- Geraads 2011.
- Yuko Hallett 2018.
- Geraads 2011.
- Yuko Hallett 2018.
- Aulagnier 2008 ; Aulagnier et al. 2017.
- Geraads 2011.
- Sahnouni et al. 2013.
- Geraads 2011.
- Sahnouni et al. 2013.
- Amri et al. 2017.
- Rueness et al. 2011 ; Gaubert et al. 2012 ; Koepfli et al. 2015 ; Viranta et al. 2017 ; Aulagnier et al. 2017.
- Geraads 2011.
- Michel & Wengler 1993a, b ; Geraads 2011.
- Aulagnier 2008 ; Aulagnier et al. 2017.
- Sahnouni et al. 2018.
- Sahnouni 2005 ; Rabhi 2009.
- Raynal et al. 2001.
- Hublin et al. 1987, 2017 ; Richter et al. 2017
- Bouzouggar et al. 2007 ; d’Errico et al. 2009 ; Barton et al. 2009 ; Nami & Moser 2010 ; Richter et al. 2010 ; Jacobs et al. 2011; Bouzouggar & Barton 2012 ; Jacobs et al. 2012 ; Linstädter et al. 2012 ; El Hajraoui et al. 2012 ; Dibble et al. 2012 ; Scerri 2013; Campmas 2017 ; Bouzouggar et al. 2018.
- Barton et al. 2016.
- Barton et al. 2005 ; Bouzouggar et al. 2008 ; Humphrey et al. 2012 ; Barton et al. 2013 ; Sari 2014 ; Sari & Kim 2017.
- van de Loosdrecht et al. 2018.
- Linstädter 2008 ; Dachy et al. 2018.
- Raynal et al. 2001.
- Daujeard et al. 2012.
- Daujeard et al. 2016.
- Daujeard et al. 2016.
- Daujeard et al. 2012.
- Bernoussi 1997.
- Raynal et al. 1993 ; Daujeard et al. 2012, 2020 ; Raynal & Mohib 2016.
- Daujeard et al. 2012, 2020.
- Daujeard et al. 2012.
- Nespoulet et al. 2008.
- Michel et al. 2009, 2010; Campmas 2012; Campmas et al. 2015, 2017.
- Michel et al. 2010.
- Campmas 2012 ; Campmas et al. 2015, 2017.
- Bougariane et al. 2010 ; Daujeard et al. 2011.
- Daujeard et al. 2011.
- Bougariane et al. 2010.
- Bougariane et al. 2010, 2012.
- Rueness et al. 2011 ; Gaubert et al. 2012 ; Koepfli et al. 2015 ; Viranta et al. 2017 ; Aulagnier et al. 2017.
- Daujeard et al. 2011.
- Aouraghe 2001; Monchot & Aouraghe 2009.
- Aouraghe 2000.
- Aouraghe 2004.
- Aouraghe et al. 2012.
- Monchot & Aouraghe 2009.
- Lenoble 2010, 2011.
- Campmas et al. 2015.
- Wrinn 2001.
- Wrinn 2001.
- Aldeias et al. 2014 ; Jacobs et al. 2011.
- Dibble et al. 2012, 2013.
- Yuko Hallett 2018.
- Yuko Hallett 2018.
- Merzoug & Sari 2008.
- Merzoug 2005.
- Geraads 2011 ; Daujeard et al. 2012.
- Amri et al. 2017.
- Merzoug 2005 ; Merzoug & Sari 2008.
- Aouraghe 2000, 2001 ; Monchot & Aouraghe 2009 ; Michel et al. 2009, 2010 ; Campmas et al. 2015 ; Yuko Hallett 2018.
- Rueness et al. 2011.
- Gaubert et al. 2012.
- Koepfli et al. 2015 ; Waters et al. 2015 ; Urios et al. 2015 ; Viranta et al. 2017 ; Aulagnier et al. 2017.
- Ferguson 1981 ;Waters et al. 2015 ; Viranta et al. 2017.
- Daujeard et al. 2012.
- Michel & Wengler 1993a, b.
- Wrinn 2001 ; Monchot & Aouraghe 2009 ; Bougariane et al. 2010 ; Daujeard et al. 2011.
- Campmas et al. 2017.
- Yuko Hallett 2018.
- Aulagnier et al. 2017.
- Daujeard et al. 2012 ; Campmas et al. 2017.
- Campmas et al. 2017.
- Michel & Wengler 1993a, b.
- Ruhlmann 1951.
- Ruhlmann 1936.
- Amani 1991.
- Ouchaou et al. 1998.
- Daujeard et al. 2011.
- Campmas et al. 2017.
- Daujeard et al. 2012.
- Yuko Hallett 2018.
- Merzoug 2005.
- Ouchaou & Amani 2002.
- Stoetzel et al. 2012.
- Aouraghe et al. 2010.
- Esperandieu et al. 1994.
- Ouchaou & Amani 2002.
- Aouraghe et al. 2010.
- Valla 1975 ; Vigne & Guilaine 2004 ; Maher et al. 2011.
- Lemjidi et al. 2018.
- Simoneau 1977 ; Rodrigue 1993 ; Lemjidi 2018.
- van Albada & van Albada 2000.
- Boube-Picot 1969.
- Limane et al. 1998.
- Esperandieu et al. 1994.
- Michel & Wengler 1993 ; Monchot & Aouraghe 2009.
- Campmas et al. 2017.
- Daujeard et al. 2012.
- Daujeard et al. 2011.
- Yuko Hallett 2018.
- Castel 2004 ; Fosse et al. 2004, 2012 ; Campmas & Beauval 2008 ; Mallye et al. 2012 ; Arilla et al. 2019.
- Campmas et al. 2018 ; Fourvel et al. 2018.
