




Introduction et problèmatique
Les castra ont fait l’objet de synthèses variées au cours de ces trente dernières années1 dont certaines se sont intéresséesnotamment à la question de l’alimentation carnée. Ainsi, les tables seigneuriales médiévales sont connues pour leurs démonstrations de richesse et d’opulence avec des mets rares et raffinés comme par exemple le héron, la cigogne ou la perdrix2. Pour autant, les études archéozoologiques menées sur ce type de contexte démontrent que ces populations aristocratiques basaient l’essentiel de leur consommation carnée sur des espèces plus “classiques”, et notamment sur la triade domestique et les animaux de basse-cour3 dont certains pouvaient être présents à l’intérieur des enceintes castrales. Dans ces résidences aristocratiques évoluaient également d’autres espèces animales tel que le chien, quasi omniprésent dans la vie quotidienne des hommes4 et qui contribue, depuis déjà longtemps, au nettoyage des rues par son rôle “d’éboueur”5. C’est ainsi qu’il laisse fréquemment les témoignages de son passage par des marques de dents sur les ossements des différentes espèces qu’il consomme. La place de cet animal est cependant bien plus variée et importante que ce simple rôle de nettoyeur : il peut être à la fois compagnon domestique, gardien des troupeaux mais aussi un allié lors des parties de chasse6. Cette proximité entre l’homme et le chien se traduit au Moyen Âge par de nombreuses représentations iconographiques sur des supports variés7.
Peu d’études archéozoologiques documentent les castra de l’Aquitaine médiévale8. Le site du Castéra, castrum primitif de Langoiran (Gironde, France), fouillé entre 2004 et 20179 vient combler ce manque en apportant des éléments de comparaison qui permettent d’amorcer la caractérisation du monde aristocratique du castrum. L’analyse des vestiges fauniques du castrum de Langoiran réalisée, s’inscrit dans une problématique générale visant notamment à comprendre le fonctionnement du castrum et à caractériser les habitudes alimentaires de ses occupants. Cette étude archéozoologique détaillée sera prochainement publiée dans le cadre de la monographie du site actuellement en préparation10. Durant cette analyse, l’identification de nombreuses traces de manducation et/ou de digestion sur les vestiges osseux, a soulevé d’autres questionnements. Dans le cadre du présent article, nos recherches se sont alors orientées sur l’identification du ou des auteur(s) de ces traces, leur(s) impact(s) sur le corpus faunique retrouvé et la place que tiennent ces “modificateurs d’ossements” au sein du groupe aristocratique du castrum.
Présentation du site
La commune girondine de Langoiran abrite aujourd’hui encore les imposants vestiges d’une forteresse de la fin du Moyen Âge (Fig. 1). Ce château, siège d’une petite seigneurie de l’Entre-deux-Mers bordelais, s’est installé à la fin du XIIIe ou au tout début du XIVe siècle11 sur les coteaux dominant la rive droite de la basse vallée de la Garonne. Il surplombe un site fossoyé de taille beaucoup plus modeste installé dans la plaine alluviale au lieu-dit le Castéra (Fig. 2). Découvert par photographie aérienne en 1985, le site du Castéra a été rapidement interprété comme l’emplacement primitif du castrum de Langoiran dont les sources écrites attestent l’existence dès le XIIe siècle12. L’exceptionnelle lisibilité du plan du site et son implantation inhabituelle dans les terres basses de la vallée de la Garonne ont justifié le lancement d’un projet de fouilles programmées dès 200413. La confirmation de la nature du site, la détermination de la chronologie et des raisons de son implantation en zone de palus, puis de sa désertion, sont les axes principaux de la réflexion menée sur le site.

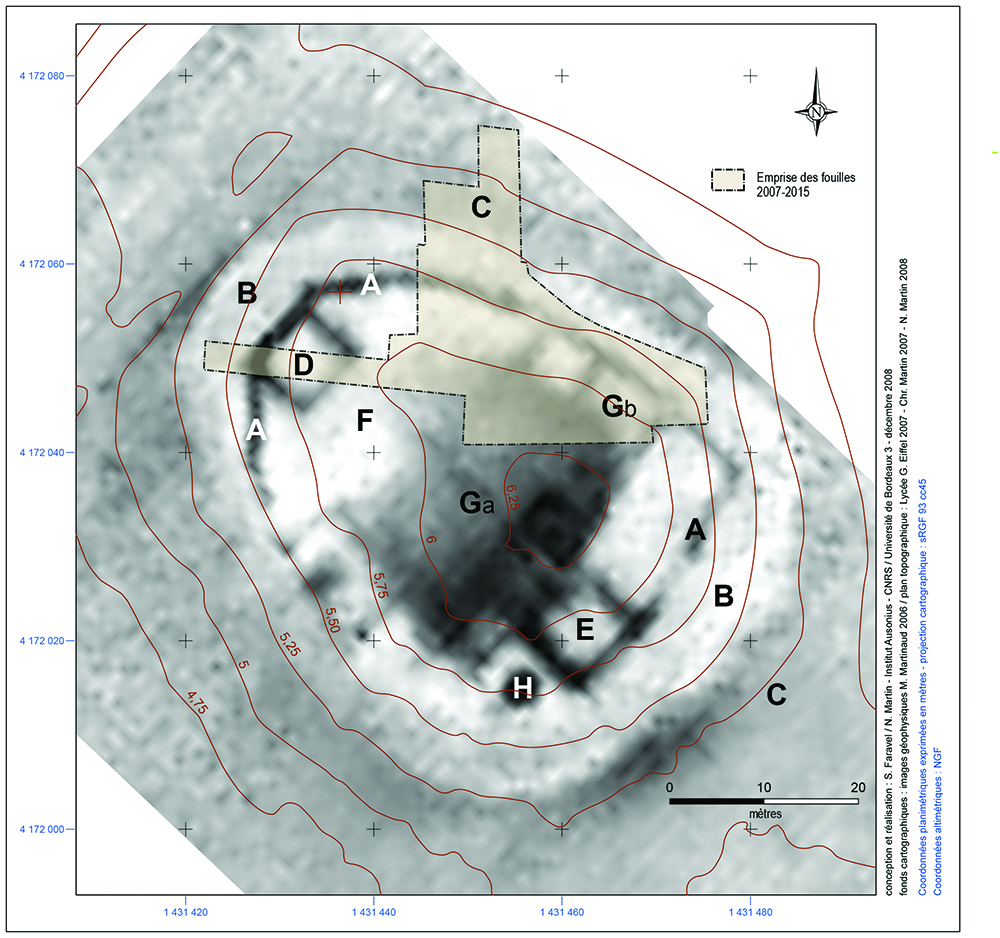
Les relevés topographiques et les prospections géophysiques réalisés en 2005 et 2007 ont confirmé et nettement complété les informations tirées des photographies aériennes (Fig. 3)14. Leurs données cumulées font apparaître clairement le plan du système défensif. Une enceinte polygonale (A), dont le tracé est inégalement conservé, est installée sur une large levée de terre de plan ovoïdal (B) et protégée par un fossé (C) assez peu lisible. Le mur d’enceinte dispose de fondations moins puissantes que les deux bâtiments carrés (D et E), probables tours appuyées sur la courtine à l’ouest et à l’est. L’espace enclos par le mur d’enceinte présente une surface d’environ 1 500 m2 ; il se partage entre, à l’ouest, une cour (F) et, à l’est, une série de constructions (G) accolées ou non au mur d’enceinte. On retrouve en (H) une structure circulaire non identifiée qui pourrait être liée à la destruction du mur d’enceinte dans cette zone. Les campagnes de fouilles réalisées entre 2007 et 201515 sur le quart nord-est du site ont permis de confirmer l’exceptionnelle précision des données fournies par la prospection géophysique tout en permettant de définir et de dater les phases d’aménagement et les modes de construction mis en œuvre. Les fouilles ont permis de comprendre la complexité apparente du bâti à l’intérieur de l’enclos fortifié en distinguant un premier état où les constructions étaient installées au cœur de l’enclos (Ga) et un second état où elles se concentrèrent en périphérie (Gb) après remblaiement de la plateforme (zone en gris foncé sur la Fig. 3).
La datation de l’ensemble soutient l’hypothèse d’une chronologie d’occupation réduite entre le XIe et le XIIIe siècles, ce qui confirme bien l’immédiate antériorité du site par rapport au château actuel. La question de l’identification du site avec le premier castrum de Langoiran semble par ailleurs acquise16. La morphologie du site, une enceinte fossoyée à l’origine, “emmottée” par la suite, permet de le placer dans la typologie régionale des fortifications de terre même si peu d’entre elles ont été fouillées jusqu’à présent17.
La typologie du mobilier trouvé18 s’inscrit dans les corpus observés sur d’autres sites de résidences aristocratiques médiévales comme ceux précédemment cités ou encore l’Isle-Bouzon dans le Gers19 et à un moindre niveau Andone en Charente20. Les objets liés à l’équipement du cheval, à la guerre ou à la chasse (éperons, fers à chevaux, carreaux d’arbalètes, trompes d’appel) ou encore au jeu21 renvoient clairement à ce contexte (Fig. 4). La proximité de la Garonne rappelle, tout comme la présence d’ichtyofaune et surtout de nombreux lests de filets et autres plombs de pêche, le rôle que le fleuve a dû tenir dans l’implantation du site et dans le développement de la seigneurie de Langoiran22.

Matériel et méthode
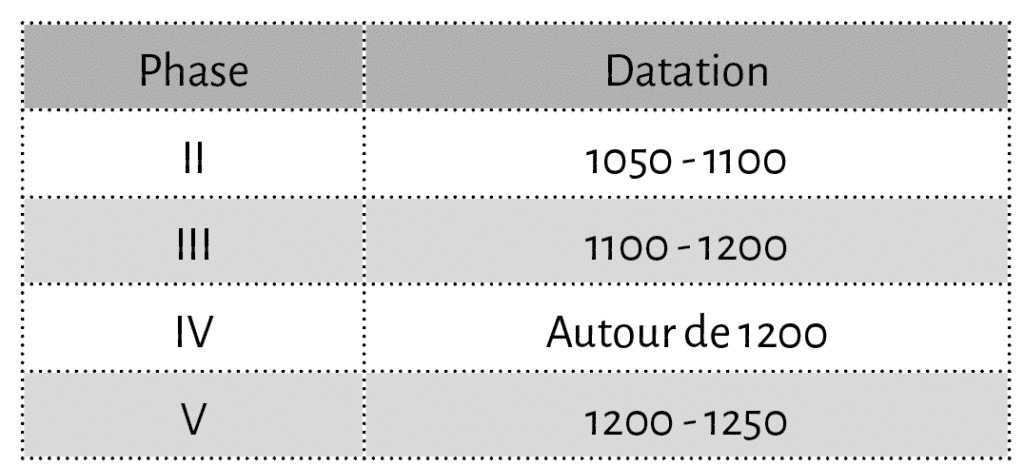
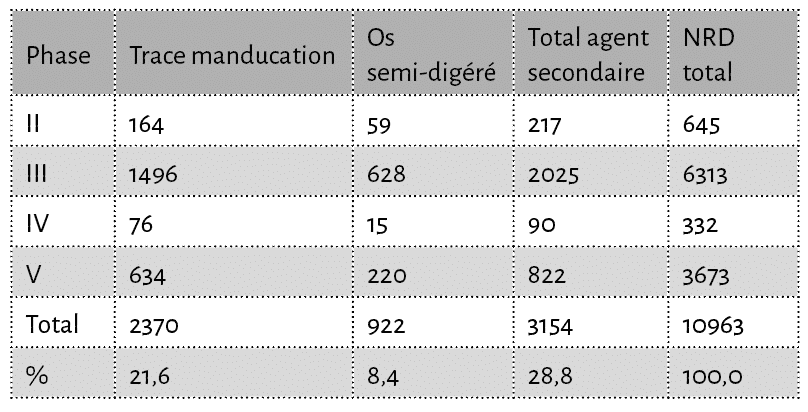
L’étude des restes fauniques de Langoiran concerne un échantillon de près de 11 000 ossements issus de plus de 150 US23, constituant les 4 phases majeures du site allant du XIe au XIIIe siècle (Tabl. 1) et réparties sur 8 secteurs distincts24. Il s’agit de la totalité des ensembles osseux et dentaires découverts lors des différentes campagnes de fouilles, à l’exception des restes de micromammifères, de poissons et de malacofaune qui font l’objet d’études séparées25. Tout le matériel a été récolté manuellement et enregistré par carré et par décapage au sein de chacune des US définies.
Chaque élément osseux a été analysé et enregistré individuellement, suivant une méthode désormais bien établie prenant en considération des données d’ordre taxinomique et taphonomique ainsi que les différentes traces humaines et animales26. Chaque reste osseux a ensuite été déterminé le plus précisément possible, tant d’un point de vue anatomique que taxinomique, grâce à la collection d’anatomie comparée du laboratoire PACEA. Il a alors été possible de dresser un spectre faunique établi à partir du nombre de restes déterminés anatomiquement et spécifiquement (NRD) et du nombre minimal d’individus (NMI)27. Lorsque la détermination spécifique n’a pu être réalisée, les vestiges osseux ont été regroupés par classe de poids de l’animal, puis dénombrés : Classe 1 (< 20 kg), Classe 2 (20-80 kg), Classe 3 (80-250 kg), Classe 4 (> 250 kg)28. Le terme “capriné” regroupe ici les restes de mouton et de chèvre pour lesquels la distinction spécifique n’a pas été possible (eg. restes fragmentaires de vertèbres, côtes, etc). De même, sous l’appellation “porc”, sont éventuellement intégrés des restes de sanglier qui n’auraient pas été identifiés spécifiquement.
Les diverses traces de boucherie et de cuisson ainsi que les traces de dents animales et de digestion, ont été identifiées grâce aux caractéristiques publiées pour les différents types de traces29 et leur localisation sur l’os, enregistrée. Les critères d’identification publiés par d’Errico et Villa30 ont été utilisés pour identifier les os semi-digérés ; ainsi l’attaque des os par les sucs gastriques se caractérise par l’apparition de perforations, un élargissement des foramens nourriciers, un émoussé des bords de fracture et un poli général de l’os. La surface corticale présente souvent des petites dépressions en forme de croissant de lune. Pour les os spongieux, nous avons utilisé le critère de dissolution de l’os cortical pour déterminer si un os était semi-digéré. Enfin, la plus grande longueur des os semi-digérés a été enregistrée.
Associés aux éléments fauniques, 16 coprolithes ont été mis au jour dans les mêmes US, dont 14 sont issus de la phase III. Pour tenter d’identifier le ou les auteur(s) de ces fèces et par la même occasion déterminer le contenu du bol alimentaire, des analyses génétiques ont été réalisées dans le laboratoire d’ADN ancien du Musée de l’Homme (J.-M. E.). Les analyses ont porté sur trois coprolithes issus de trois US différentes de la phase III en recherchant l’ADN mitochondrial de Carnivore (Canis) et de Suidé (Sus). Pour cela, différents couples d’amorces ont été utilisés pour amplifier l’ADN. Les amorces de Canidé correspondent à des régions conservées dans cette famille, mais encadrent une région suffisamment variable de la région de contrôle du génome mitochondrial pour permettre l’identification de l’espèce. Les amorces de Suidé ciblent le gène mitochondrial cytB. Deux couples d’amorces ont été utilisés, prédisant respectivement l’amplification de fragments de 85 et 102 paires de bases. Les ADN amplifiés ont été séquencés pour vérifier la taille des fragments et la présence d’endommagements attendus pour l’ADN ancien.
Résultats
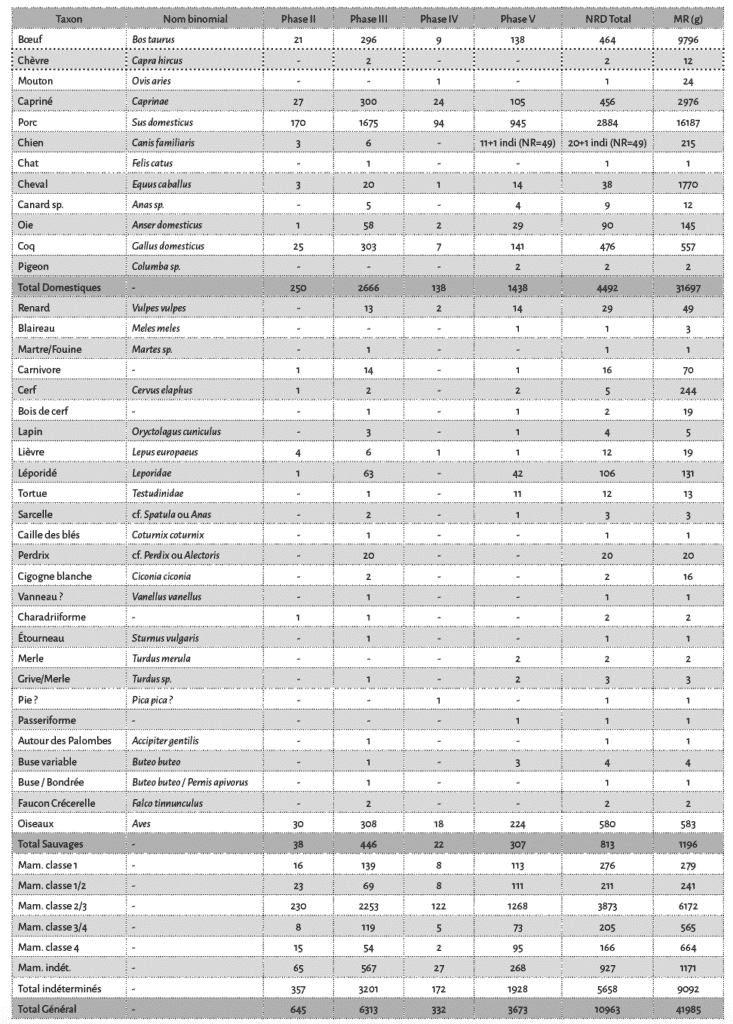
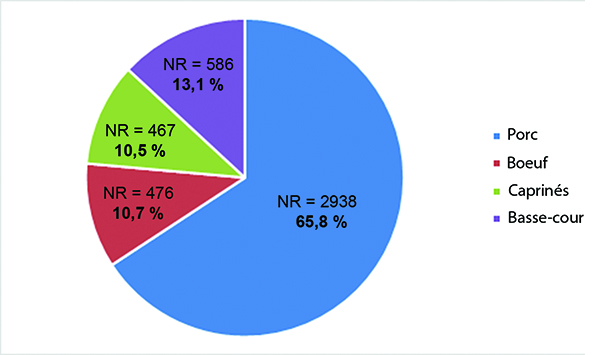
Près d’une quarantaine de taxons ont été inventoriés bien que la moitié (52 %) des restes n’aient pu être déterminés au rang de l’espèce. Les phases III et V, qui représentent les deux grandes phases d’occupation du site, concentrent la plus grande diversité animale (Tab. 2). La triade domestique domine l’assemblage osseux dans toutes les phases chronologiques. À l’échelle du site, le porc représente les trois quarts des restes de la triade, le bœuf et les caprinés étant ensuite représentés en parts égales. De plus, 5 espèces de carnivores ont été reconnues et la part de l’avifaune est elle aussi importante avec 16 taxons identifiés.
La faune
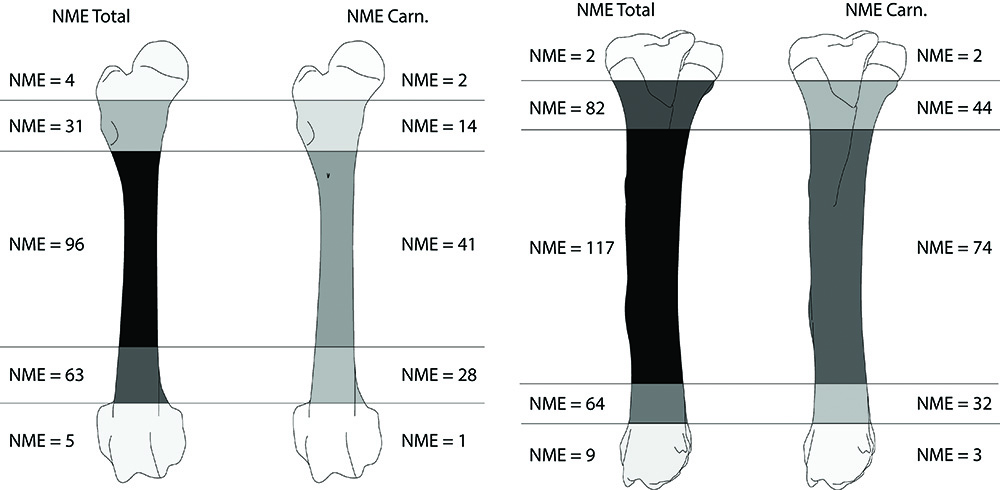
En ce qui concerne la faune domestique, dans toutes les phases chronologiques le porc domine largement l’assemblage faunique, tant en nombre de restes déterminés qu’en masse des restes (Fig. 5 ; Tab. 2). Avec un total de 2884 pièces osseuses, le porc représente un peu plus de la moitié des restes déterminés et un peu plus de 20 % des vestiges sont porteurs de traces anthropogéniques. Les autres membres de la triade sont identifiés dans des proportions identiques avec 454 et 459 pièces respectivement pour le bœuf et les caprinés. Sur ces taxons, les traces anthropogéniques sont présentes à hauteur de 6 à 10 %. Le cheval est identifié à partir de 38 pièces dont 19 sont des dents isolées et parmi celles-ci, 9 sont des déciduales de chute impliquant la présence des poulains dans l’enceinte. Des traces de boucherie sont observées sur 3 ossements. Le chien totalise 69 vestiges dont 49 appartiennent à un seul et même individu adulte et au moins 2 autres individus sont dénombrés sur la base des combinaisons des portions d’humérus. L’ensemble de ces restes est dépourvu de traces d’origine anthropique. Le dernier animal domestique identifié est le chat, représenté par un humérus. Les membres de la basse-cour sont représentés ici par 577 restes dont une majorité (82 %) renvoie au coq, ainsi qu’à l’oie et dans une moindre mesure au canard et au pigeon. Les traces d’exploitation de la viande sont observées sur 6 % des restes de ces espèces.
De nombreuses espèces sauvages ont été identifiées dont des petits carnivores, du gibier à poil et à plume (Tab. 2). Les carnivores sont essentiellement représentés par le renard, avec 29 restes. Il faut également ajouter une ulna de blaireau et une ulna de martre ou de fouine. Le gibier à poil est représenté par le cerf et les léporidés. La présence du cerf est documentée par deux fragments de bois qui ont été manufacturés31 et des fragments d’un humérus, d’un radius, de deux tibias et d’un métatarsien. Les léporidés sont représentés par 122 restes osseux dont 4 attribués spécifiquement au lapin et 12 au lièvre. Le spectre aviaire est diversifié avec une douzaine de taxons identifiés. Certaines espèces ont été exploitées : un reste de cigogne et un autre de buse variable présentent des stries de découpe.
Aparté sur le chien
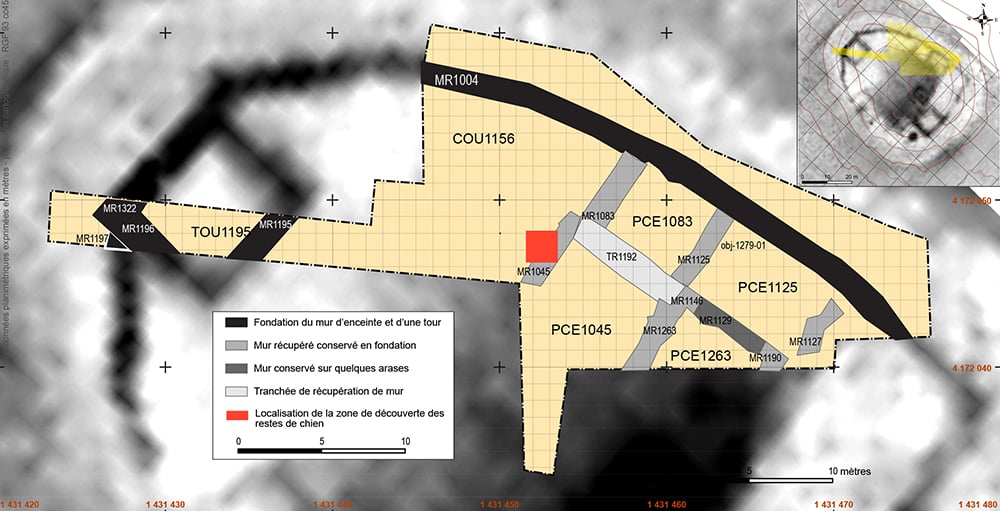
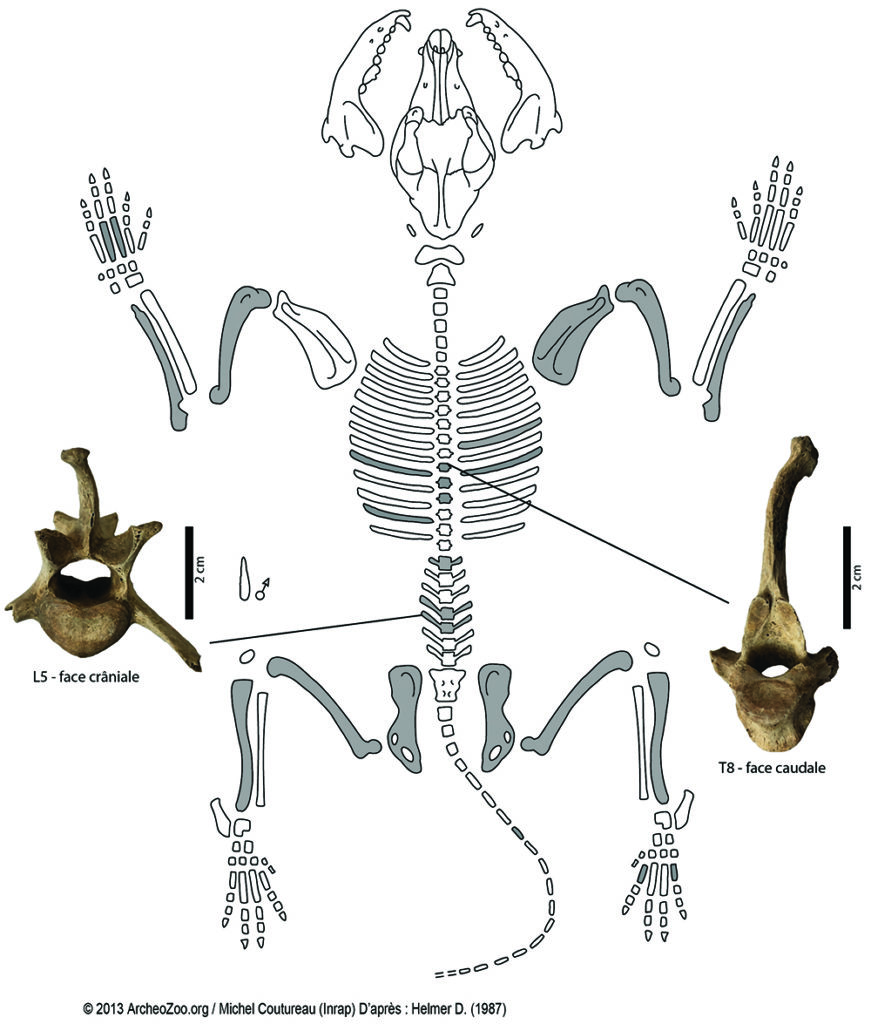
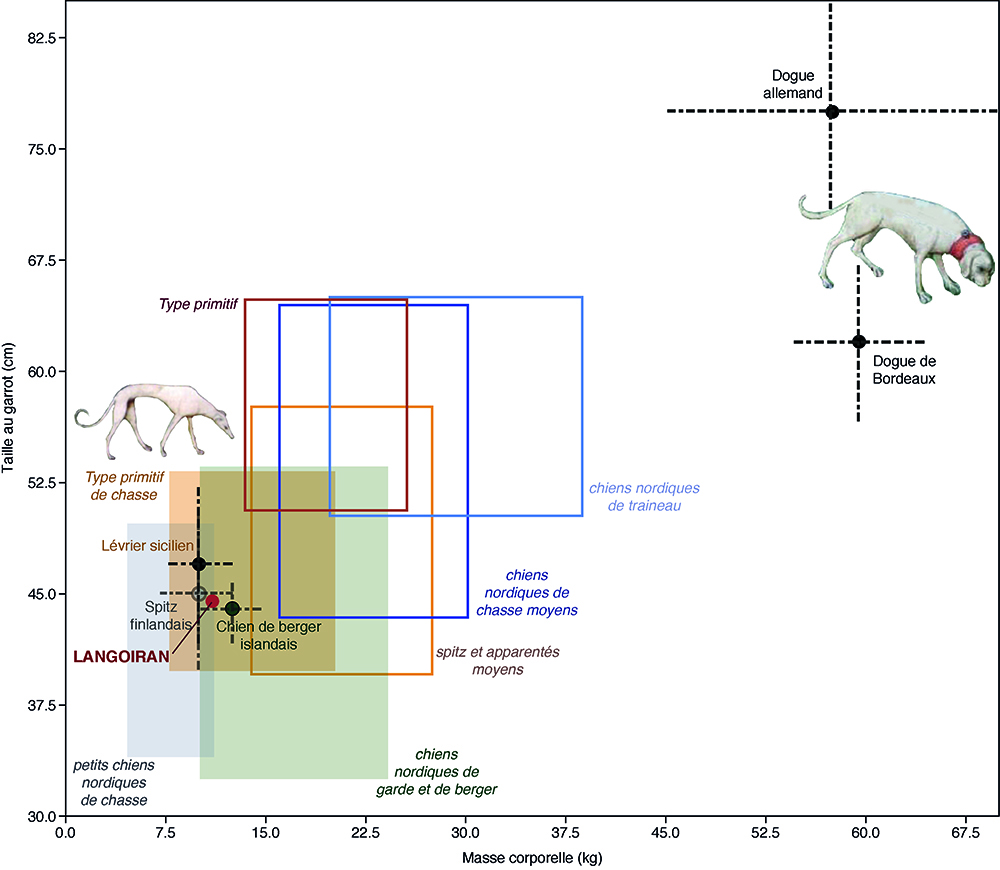
À Langoiran, cet animal semble disposer d’un statut particulier. En effet ces restes sont en majorité complets et ne portent aucune trace anthropogénique ni de traces d’un agent secondaire. Les 69 restes identifiés renvoient à 3 individus et deux gabarits différents semblent se dégager avec deux chiens de petite taille et un chien plus gros. Les longueurs prises sur les métatarsiens 2 et 5 du chien partiellement complet sont respectivement de 50 et 51 mm. Ces mesures ne sont pas compatibles avec celle du quatrième métatarsien isolé dont la longueur atteint 75 mm ; une différence de l’ordre de 10 % étant généralement observée chez les canidés entre la longueur de ces métapodiens. Les restes appartenant au même individu ont été découverts dans l’US 1039, au niveau du mur MR104532, dans les carrés A98 (5), A98/99 (5), B98 (32) et B99 (7) (Fig. 6). Ils se réfèrent à l’ensemble du squelette à l’exception du crâne (Fig. 7). Il s’agit d’un chien ostéologiquement mature, de petit gabarit et de sexe indéterminé. Sa stature au garrot peut être estimée à environ 44,5 cm selon les coefficients de Harcourt33, à partir des longueurs du fémur (TL = 144,4 mm) et du tibia (TL = 150,3 mm). Sa masse corporelle est évaluée à environ 11 kg, selon les équations établies sur des loups34. À titre de comparaison, si l’on ne tient compte que des races de chiens appartenant au groupe de type Spitz et de type primitif (groupe 5), ces proportions sont comparables à celles du lévrier sicilien et ne sont donc pas sans rappeler le canidé figuré sur le pion de jeu découvert dans le castrum35. En effet, en considérant la stature et la masse corporelle estimées pour le chien de Langoiran, ainsi que les types de chiens décrits à cette époque, notre individu devait clairement plus se rapprocher d’un lévrier que d’un molosse (Fig. 8). Il s’inscrit dans les gabarits du lévrier sicilien et des petits chiens nordiques de chasse ou de berger mais ces derniers n’étaient pas présents dans notre région au Moyen Âge. La présence de ce type de chien dans le castrum pourrait par ailleurs être liée à son aptitude à la chasse, tout comme les lévriers découverts à Noyon36. D’un point de vue paléo-pathologique, plusieurs éléments du squelette axial de cet animal sont remarquables. Les processus épineux de plusieurs des vertèbres identifiées présentent une courbure latérale. Sur une thoracique (identifiée de rang 8) et sur une lombaire (L5), cette courbure est nettement marquée vers la droite (Fig. 7) Eu égard aux travaux menés sur ce type d’anomalies37, il serait présomptueux de déterminer les causes de ces déviations sans pousser l’analyse à l’échelle de la structure interne de l’os. Nous pouvons toutefois rappeler qu’elles peuvent être liées à des anomalies structurelles, à une pathologie, ou encore aux conditions de vie de l’animal, et peuvent donc faire écho au statut de ce chien au sein du castrum. L’US 1039, qui renferme ces ossements, correspond à un niveau de remblai qui vient combler la tranchée de récupération du mur M1045 détruit en phase IV. Ce mur marquait la limite entre l’espace bâti, à l’est, et la cour COU1156, à l’ouest (Fig. 6). Le cadavre de ce chien a donc pu être initialement déposé puis recouvert, dans la cour, au pied du mur MR1045. Au moment de la récupération du mur, l’organisation des restes du chien a été bouleversée et une partie de ces éléments se sont retrouvés ensuite dans le comblement de la tranchée de récupération. Ce traitement pourrait alors être interprété comme une volonté de donner une sépulture à cet individu, ou simplement d’enterrer un animal mort en le mettant à l’abri d’éventuels charognards.
Les traces d’agents secondaires
Les traces de manducation et de digestion
Le point remarquable de cette série réside dans le fait que de nombreux restes portent des traces de manducation et/ou de digestion. Ainsi, un total de 3154 restes osseux, soit 28,8 % du nombre de restes total, présentent des marques de consommation (enfoncements, perforations, sillons, mâchonnement, encoches, semi-digérés) dont 922 (soit 8,4 %) sont semi-digérés (Tab. 3). Chez le porc un quart des restes présentent des traces de manducation alors que chez le bœuf ces traces sont observées sur près de 6 % des restes et sur 8 % des restes de caprinés. Enfin, pour le cheval le nombre d’éléments affectés par des traces de dents est de 14 restes sur les 39 déterminés.
Ces traces de manducation se déclinent en des enfoncements et perforations circulaires ou allongées de la surface corticale, des sillons marqués, des cylindres osseux mâchonnés ou des encoches de perforation38 (Fig. 9 et 10). D’autres types de traces ont été identifiés. Il s’agit d’enfoncements en double rangée parallèle et régulièrement alignés présents sur les faces externes des ossements (Fig. 11).



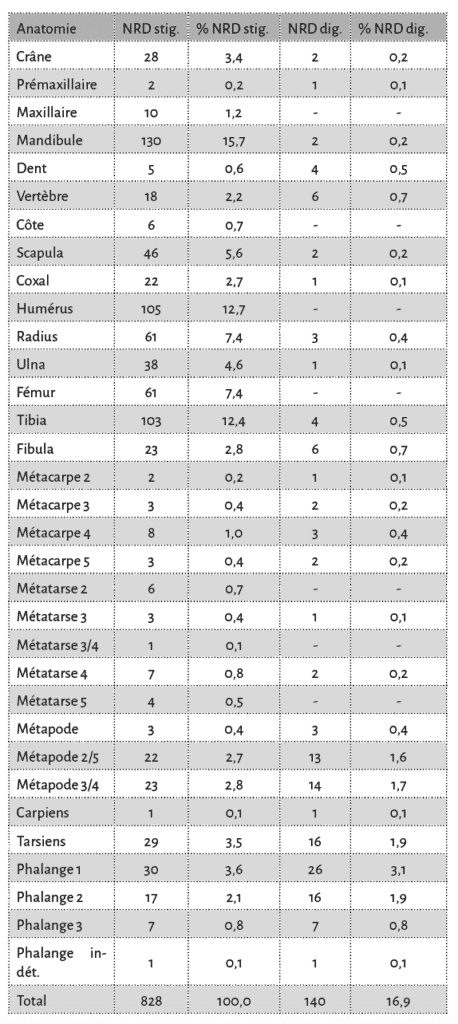
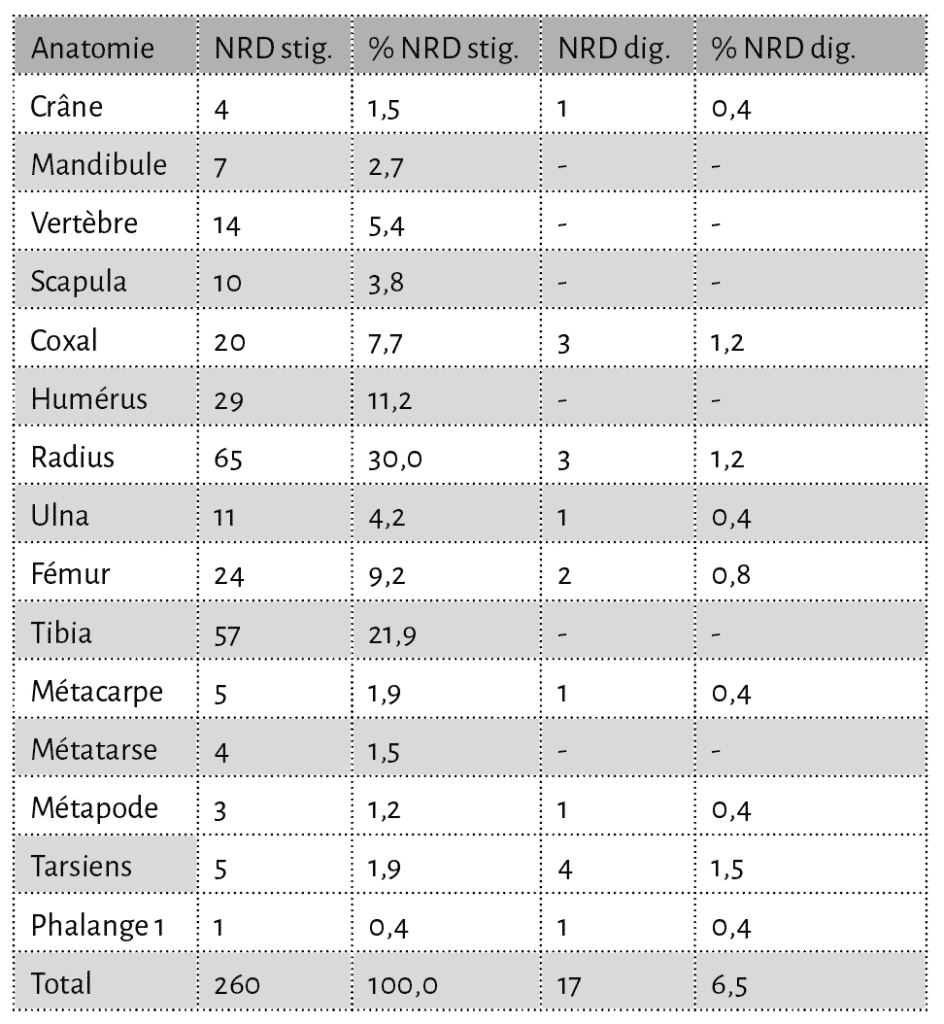
Toutes les portions squelettiques du porc et des caprinés sont affectées par des traces d’action animale (Tab. 4 et 5). Lorsque l’on s’intéresse à la séquence de consommation des os longs du porc (taxon le plus abondant), on constate rapidement que ces ossements sont attaqués de façon “classique” ; ce sont les extrémités des os qui sont préférentiellement consommées39 (Fig. 12 et 13). En effet, les décomptes établis, en nombre minimal d’éléments pour les os des membres, montrent que les extrémités articulaires de ces os ont pratiquement toutes disparu. Seuls les fragments diaphysaires sont trouvés en quantité et portent pour la grande majorité des traces de manducation.
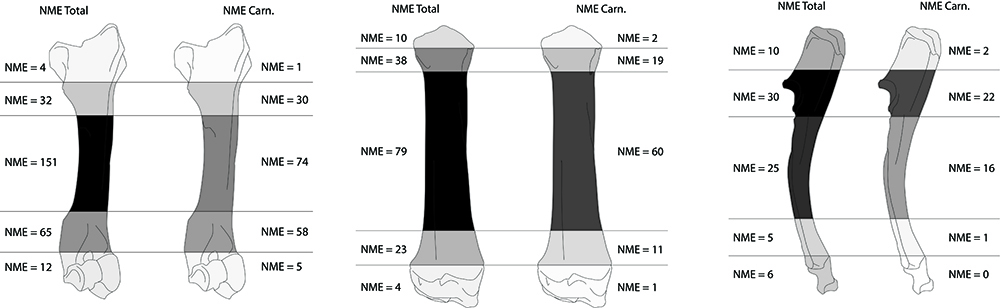
La distribution de la taille des vestiges fauniques suit la loi de Poisson, de même que celle des ossements modifiés secondairement (Fig. 14).
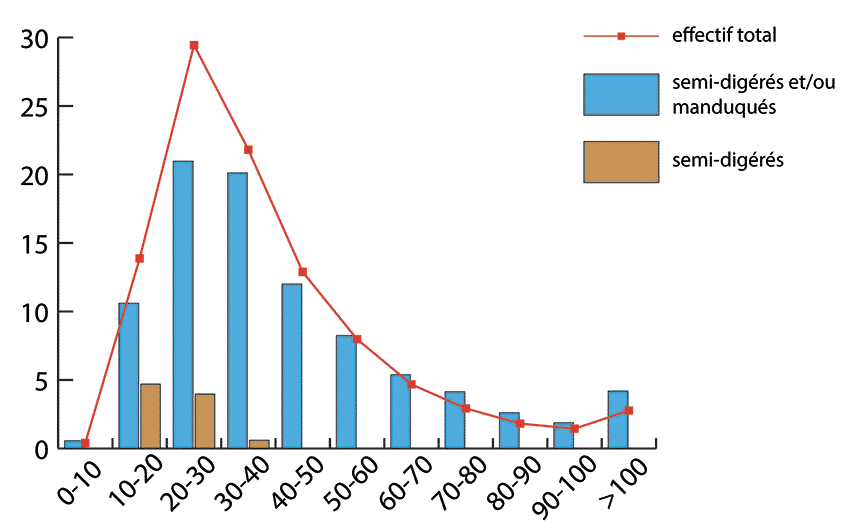
En ce qui concerne les ossements semi-digérés, leur longueur se distribue préférentiellement sur des valeurs basses avec des tailles pouvant atteindre au maximum 50 mm. Cependant, il faut noter que la grande majorité d’entre eux (93 %) ont une taille comprise entre 10 et 30 mm (Fig. 14 et 15). Enfin certains éléments (4,5 %), correspondant principalement aux os des extrémités des pattes, sont complets (Fig. 15). Ces éléments semi-digérés se rapportent majoritairement à des éléments du squelette axial (35 %), à des os courts des pattes (30 %), à des os longs (19 %), à des éléments de ceintures (10 %) et des éléments crâniens (6 %). Ainsi, toutes les portions du squelette sont touchées par ce type d’attaque. Enfin, sur l’ensemble du matériel osseux, 462 éléments, soit 4,2 % du NR, portent à la fois des stigmates anthropiques et des traces de manducation.

Les coprolithes
Parmi les 16 coprolithes identifiés, 3 ont fait l’objet d’un prélèvement pour analyse ADN. De l’ADN de canidé (Canis lupus ssp.) et de suidé (Sus scrofa) a été détecté dans ces trois coprolithes, et identifié formellement en établissant la séquence des fragments amplifiés. La taille des fragments amplifiés est de 89 paires de bases pour Canis lupus/familiaris.
Cependant, des différences d’un échantillon à l’autre peuvent être notées. Pour deux coprolithes (n° d’inventaire 1056 et 1058) le taux de succès des tentatives d’amplification est faible (33 % de réussite), indiquant que l’ADN est peu abondant car probablement dégradé. Pour le troisième coprolithe (n° d’inventaire 1025), on note en revanche 100 % de réussite (Canis lupus/familiaris : 3/3 ; Sus scrofa : 6/6) pour les amplifications par PCR. Ceci indique pour ce coprolithe une bonne préservation de l’ADN, qui présente tout de même sur une partie des fragments les substitutions attendues (CG/TA) pour un ADN ancien authentique. Ainsi, quel que soit le degré de préservation du matériel génétique, on trouve dans chacun des trois coprolithes de l’ADN de canidé et de suidé.
Discussion générale
Les traces d’agents secondaires
Près de 29 % du NRD montrent des traces de consommation par un agent secondaire. De telles proportions d’éléments exploités indiquent que ces agents ont eu un accès immédiat aux ossements. Dans ce contexte du castrum, il est facile d’imaginer que ces animaux ont hérité des déchets de préparation bouchère et des déchets d’assiettes40. Cette bonne gestion des rebuts domestiques permettait à la fois de nourrir facilement les animaux mais aussi d’éliminer une partie des détritus gênants pour des questions évidentes d’hygiène.
Se pose alors la question du ou des agent(s) responsable(s) de ces transformations ? L’identification de l’agent destructeur n’est pas toujours aisée et il est souvent difficile de déterminer s’il s’agit, dans un contexte tel que celui du Castéra, de l’action des chiens et/ou des porcs. Néanmoins, certains vestiges présentent des caractéristiques permettant d’attribuer l’intervention sur l’os à un agent précis.
La distribution de la taille des ossements modifiés secondairement est comparable à celle des vestiges fauniques identifiés à Langoiran ce qui démontre qu’il n’y a pas de sélection, par les agents secondaires, des restes qu’ils ont consommés. Les mêmes conclusions peuvent être tirées à partir de l’étude des restes semi-digérés ; toutes les portions anatomiques sont identifiées parmi ces restes. Tous les déchets de boucherie étaient jetés dans l’enceinte et par là même, rendus accessibles à un ou des agents modificateurs secondaires.
Si l’on se réfère maintenant à la morphologie des traces présentes sur les surfaces osseuses, la plupart se rapproche de ce qui est décrit suite à la consommation et la modification d’ossements par des canidés (enfoncements et perforations circulaires, encoches de fracturation, sillons, etc.). Cependant, la position alignée et symétrique de certains enfoncements se démarque (Fig. 11). Ces traces, associées à de légers sillons sur les angles des diaphyses, sont assez semblables à celles décrites suite à la consommation d’os par les suidés41. Le porc a donc également pu avoir accès aux différents ossements dont ceux de ses congénères, ce qui implique leur présence dans l’enclos fortifié. Leurs capacités de “nettoyeur” tout comme celles du chien, confèrent à ces espèces un atout majeur pour être acceptés dans l’enceinte du castrum, permettant ainsi d’éliminer une partie des déchets tout en fournissant de la nourriture à moindre frais.
En ce qui concerne les os semi-digérés, leur taille est comparable avec les données publiées dans les différents travaux qui concernent les études coproscopiques entreprises sur les loups42. Des résultats similaires produits par les dholes ont été observés, notamment sur le site de la grotte du Noisetier43. À notre connaissance, bien que plusieurs travaux existent sur la consommation d’ossements par les suidés44, il n’existe pas d’étude détaillée concernant les os semi-digérés par ces animaux. Il ne nous est donc pas possible de savoir si, au moins une partie des os semi-digérés de Langoiran, pourrait résulter d’une consommation par les porcs.
En outre, rappelons que de l’ADN de canidé et de suidé a formellement été identifié dans les coprolithes analysés, suggérant que l’un et l’autre ont joué un rôle de consommateur ou d’espèce consommée. Les ossements de chiens ne portent aucune trace de consommation et le nombre de leurs restes est relativement faible par rapport au NRD total (0,6 % du NRD ou 0,2 % du NRD si l’on fait exception de l’individu partiel). Les cadavres de chiens ne sont donc pas éliminés dans les “poubelles” des habitants du castrum. Rappelons par ailleurs que les restes d’un individu quasi complet ont été découverts. En comparaison, le NRD du porc est de 2918 restes, soit 54,1 % du NRD. La proportion de porc étant 90 fois supérieure à celle du chien, il est fort probable que ce soit le chien qui ait préférentiellement consommé des restes de porcs, plutôt que le porc de rares restes de chiens, et qu’il soit ainsi le principal producteur de ces coprolithes.
Néanmoins, cette hypothèse n’a pu être totalement confirmée par les analyses génétiques des coprolithes qui n’ont recherchés ici, que la présence/absence des ADN concernés. Des analyses complémentaires sur la quantification de ces ADN et/ou sur la recherche de parasites spécifiques à ces deux espèces45 pourraient peut-être venir éclaircir cette interrogation.
Conclusion
L’analyse des restes fauniques de la fouille du Castéra a permis de montrer qu’une forte proportion de restes osseux porte les traces de l’action combinée de deux agents secondaires venus charogner les déchets de cuisine et en faire disparaitre une partie. L’analyse conjuguée de ces traces et des données paléogénomiques indiquent que le chien et le porc en sont à l’origine, ce qui nous renvoie à l’expression périgourdine “meitat chen, meitat pòrc”46 (moitié chien, moitié porc). Une telle gestion des déchets, permet à la fois de nourrir ces animaux à moindre coût mais aussi d’entretenir les espaces de vie et de maintenir l’hygiène des lieux.
Ainsi, le chien et le porc jouent un rôle important sur le site, rappelant les contacts directs qu’ils ont eu avec les hommes dans le castrum. Ces relations de proximité sont encore plus marquées pour le chien qui a une situation privilégiée sur le site. En effet, en plus de son rôle de “nettoyeur”, il n’est pas consommé par les occupants des lieux. Le chien semble être le seul animal à avoir bénéficier d’un traitement particulier après sa mort sur le site du Castéra. Ses proportions, qui se rapprochent du lévrier sicilien, rappellent fortement la représentation du pion de jeu découvert dans l’enceinte ou dans une moindre mesure les lévriers découverts dans les niveaux du XIIe siècle à Noyon. On peut ainsi considérer que, si ces chiens sont un traceur de milieu privilégié voire aristocratique, leur apport comme auxiliaire de chasse n’est peut-être pas la seule explication de la présence de ces animaux sur ce site castral et que leur place au sein de la population va bien au-delà de cette fonction. De son rôle de compagnon domestique jusqu’à ses capacités de gardiennage, l’éventail des fonctions potentielles du chien est assez large, et ce sont autant d’éléments qui ont pu motiver sa présence dans le castrum de Langoiran.
Remerciements
Cette recherche a pu être menée grâce au soutien financier du projet Labex TeHoTeCa (ANR-10-LABX-52) et du ministère de la Culture et de la Région Nouvelle Aquitaine (projet “Habiter, consommer, échanger dans l’Aquitaine garonnaise médiévale”). Nous tenons donc à remercier tous les acteurs de ces projets qui ont permis la réalisation de ce travail. Nous remercions également les laboratoires PACEA et Ausonius, qui nous ont donné accès à leurs locaux et notamment à la salle d’anatomie comparée de PACEA, ainsi qu’auPlateau Technique du MNHN, site du Musée de l’Homme (plateforme Paléogénomique et Génétique Moléculaire, Musée de l’Homme) et au Service de Systématique Moléculaire du MNHN (UMS 2700 OMSI CNRS-MNHN), où les expériences sur l’ADN des coprolithes ont été réalisées. Nos remerciements vont également à nos collègues, en particulier Véronique Laroulandie et Monica Gala, qui ont apporté leur expertise pour la détermination de certains spécimens aviaires récalcitrants et à William Rendu pour ses conseils avisés. Nous remercions enfin les relecteurs qui par leurs commentaires ont contribué à améliorer la qualité de cet article.
Bibliographie •••
- d’Anthenaise, C. (2002) : Le livre de chasse de Gaston Phébus, Paris.
- Araguas, P. et Barraud, D. (2006) : “Archéologie des résidences aristocratiques médiévales en Aquitaine”, in : Résidences aristocratiques, résidences du pouvoir entre Loire et Pyrénées, Xe-XVe siècles, Actes du colloque de Pau des 3, 4 et 5 octobre 2002, Archéologie du Midi Médiéval Suppl. 4, 13-22.
- Araguas, P. et Faravel, S. (2006) : “Les châteaux de Langoiran, premier bilan archéologique”, in : Château, livres et manuscrits IXe – XXIe siècles, Actes des Rencontres d’archéologie et d’histoire en Périgord, Périgueux 23, 24 et 25 septembre 2005, Ausonius Scripta Mediævalia, Bordeaux, 249-263.
- Audoin, F. (1986) : Ossements animaux du Moyen Âge au monastère de La Charité-sur-Loire, Paris.
- Audoin-Rouzeau, F. (1993) : “Hommes et animaux en Europe. Corpus de données archéozoologiques et historiques”, Dossier de documentation archéologique, 16, Paris.
- Baratay, É. (2003) : Et l’homme créa l’animal, Paris.
- Barja Núñez, I. et Corona-M., E. (2007) : “El análysis del excretas desde la etología y la arqueozoología. El caso del lobo ibérico”, in : E. Corona-M. et J. Arroyo-Carales, éd. 2007, Human and faunal relationships reviewed: an archaeozoological approach, BAR Int. Ser., Oxford, 113-121.
- Beck, C. (1984) : “Animal et vie quotidienne en France et en Italie d’après les vestiges ostéologiques (XIe-XVe siècles)”, in : F. Cerdan, éd. 1984, Actes des congrès de la Société des historiens médiévistes de l’enseignement supérieur public, 15e congrès, Toulouse. Le monde animal et ses représentations au Moyen-Âge (XIe – XVe siècles), Toulouse, 103-120.
- Beuchet, L. et Clavel, B. (2014) : “Les cuisines et l’alimentation dans un château breton au Moyen Âge et à l’époque moderne d’après l’archéologie : l’exemple du château du Guildo (Côtes-d’Armor) du XIe au XVIIe siècle”, in : Cocula, A.-M. et M. Combet, éd. 2014, Châteaux, cuisines & dépendances, Ausonius Scripta Mediævalia 26, Bordeaux, 107-126.
- Binford, L. R. (1981) : Bones : Ancient Men and Modern Myth, New York.
- Blancou, J. (2000) : Histoire de la surveillance et du contrôle des maladies animales transmissibles, Paris.
- Blumenschine, R. J., Marean, C. W. et Capaldo, S. D. (1996) : “Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussion marks, and carnivore tooth marks on bone surfaces”, Journal of Archaeological Science, 23, 4, 493-507.
- Bodson, L. (1979) : “Le chien. Évolution des races, de l’élevage et de l’utilisation du chien”, Ethnozootchnie, 25, 13-21.
- Borvon, A. (2012) : Acquisition des ressources animales, alimentation carnée et distinction sociale en Anjou de la fin du Xe au début du XIIe siècle. Étude archéozoologique du site de Montsoreau (Maine-et-Loire), thèse de doctorat, Université Paris I Panthéon-Sorbonne.
- Boudadi-Maligne, M. (2010) : Les Canis pléistocènes du Sud de la France : approche biosystématique, évolutive et biochronologique, thèse de doctorat, Université Bordeaux I.
- Bourgeois, L. (2009) : Une résidence des comptes d’Angoulême autour de l’an Mil – Le castrum d’Andone (Villejoubert, Charente), Caen.
- Brugal, J.-P., David, F., Farizy, C. (1994) : “Quantification d’un assemblage osseux : paramètres et tableaux”, in : M. Patou-Mathis, éd., 1994, Outillage peu élaboré en os et bois de cervidés – 6ème Table Ronde Taphonomie, Bone modification, Paris, 143-153.
- Buffière, É. (2015) : Vivre dans une résidence aristocratique au XIIIe s, Étude du petit mobilier du site du Castéra, Langoiran (33), mémoire de master 2 d’archéologie, sous la direction de Sylvie Faravel, Université Bordeaux Montaigne, 2015.
- BSR 2016 : DRAC Aquitaine – Service Régional de l’Archéologie. Bilan Scientifique 2016.
- Caillat, P. et Laborie, Y. (1997) : “Approche de l’alimentation carnée des occupants du castrum d’Auberoche (Dordogne) d’après les données de l’archéozoologie”, Archéologie du Midi médiéval, 15-16, 161-177.
- Campmas, E. et Beauval, C. (2008) : “Consommation osseuse des Carnivores : résultats de l’étude de l’exploitation de carcasses de bœufs (Bos taurus) par les loups captifs”, Annales de Paléontologie, 94, 167-186.
- Capaldo, S. D., et Blumenschine, R. J. (1994) : “A Quantitative Diagnosis of Notches Made by Hammerstone Percussion and Carnivore Gnawing on Bovid Long Bones”, American Antiquity, 59, 4, 724-748.
- Castel, J.-C. (2004) : “L’influence des canidés sur la formation des ensembles archéologiques. Caractérisation des destructions dues au loup”, Revue de Paléobiologie, 23, 2, 675-693.
- Clavel, B. (2001) : “L’animal dans l’alimentation médiévale et moderne en France du Nord (XIIe – XVIIe siècles)”, RAP Numéro spécial 19.
- Collectif BnF : “Bestiaire enluminé”, http://expositions.bnf.fr/bestiaire/index.htm
- Costamagno, S. (1999) : Stratégies de chasse et fonction des sites au Magdalénien dans le Sud de la France – Tome I et II, thèse de doctorat, Université Bordeaux I.
- Didierjean, F. (1988) : “Archéologie aérienne dans la région Aquitaine : approche méthodologique”, in : Le point sur la prospection aérienne : journée d’archéologie aérienne organisée par le Centre de recherches archéologiques à l’université de Toulouse-Le Mirail le 20 avril 1985, UA 997 CNRS, Toulouse, 45-50, fig. 3.
- Dominguez-Solera, S. D. et Dominguez-Rodrigo, M. (2009) : “A taphonomic study of bone modification and of tooth-mark patterns on long limb bone portions by suids”, International Journal of Osteoarchaeology, 19, 345-363.
- Dumolin, M. (1935) : “L’abbaye Saint-Jean de Sorde (Landes)”, Bulletin Monumental, 94, 1, 5-28.
- Durand, A., Forest, V., Ruas, M.-P. et Gardeisen, A. (1997) : “Approches bioarchéologiques de l’habitat castral languedocien. Huit sites de la bordure méridionale du Massif central (Xe-XVIe siècle)”, Histoire et Sociétés Rurales, 8, 2, 11-32.
- d’Errico, F. et Villa, P. (1997) : Holes and grooves: the contribution of microscopy and taphonomy to the problem of art origins, Journal of Human Evolution, 33, 1-31.
- Estan-Nadal, M., Cáceres, I., Fosse, P. (2010) : Characterization of current coprogenic sample originated by Canis lupus as a tool for identifying a taphonomic agent, Journal of Archaeological Science, 37, 2959-2970.
- Faravel, S., dir. (2004) : Le Castéra et le Palus de Langoiran (Gironde), rapport de prospection 2004, Service Régional de l’Archéologie d’Aquitaine, Bordeaux, UMR Ausonius, décembre 2004.
- Faravel, S. (2014) : “Le site du Castéra, commune de Langoiran (Gironde)”, in : Bourgeois, L. et Rémy, C., dir. 2014, Demeurer, défendre, paraître. Orientations récentes de l’archéologie des fortifications et des résidences aristocratiques médiévales entre Loire et Pyrénées. Colloque organisé par le CESCM et le SRA Poitou-Charentes, et la ville de Chauvigny, Chauvigny, 14-16 juin 2012, Chauvigny, 351-367.
- Faravel, S., dir. (2015) : Le castrum du Castéra (commune de Langoiran, Gironde). Rapport de fouille programmée 2014, SRA Aquitaine.
- Faravel, S., dir. (en préparation) : Le castrum du Castéra (commune de Langoiran, Gironde). Rapport de fouille programmée 2015, Bordeaux, UMR Ausonius.
- Faravel, S., dir. (en préparation) : Le castrum du Castéra (commune de Langoiran, Gironde). Monographie du site.
- Fisher, J. W. (1995) : “Bone Surface Modifications in Zooarchaeology”, Journal of Archaeological Method and Theory, 2, 1, 7-68.
- Fosse, P. (1994) : Taphonomie Paléolithique : les grands mammifères de Soleilhac (Haute-Loire) et de Lunel-Viel 1 (Hérault), thèse de doctorat, Université Aix-Marseille I.
- Fosse, P., Avery, G., Selva, N., Smietana, W., Okarma, H., Wajrak, A., Fourvel, J.-B. et Madelaine, S. (2011) : “Taphonomie comparée des os longs d’ongulés dévorés par les grands prédateurs modernes d’Europe et d’Afrique (C. lupus, P. brunnea)”, in : J.-P. Brugal, A. Gardeisen & A. Zucker, éd., 2011, Prédateur dans tous leurs états. Evolution, Biodiversité, Interactions, Mythes, Symboles. XXXIe rencontres internationales d’archéologie et d’histoire d’Antibes, APDCA, Antibes, 127-156.
- Goret, J.-F., Talon, M., Yvinec, J.-H., Derbois, M., Mouny, S. et Poplin, F. (2009) : “Le jeu d’échecs de Noyon dans son contexte archéologique et historique”, RAPSuppl. 2009, 79-119.
- Greenfield, H. J. (1988) : “Special Studies : Bone Consumption by Pigs in a Contemporary Serbian Village: Implications for the Interpretation of Prehistoric Faunal Assemblages”, Journal of Field Archaeology, 15, 473-479.
- Harcourt, R. A. (1974) : “The Dog in Prehistoric and Early Historic Britain”, Journal of Archaeological Science, 1, 151-157.
- Hesse, B. et Wapnish, P. (1985) : Animal bone archaeology. From objectives to analysis, Washington.
- Klein, R. G. et Cruz-Uribe, K. (1984) : The analysis of animal bones from archeological sites, London.
- Laborie, Y. (2006) : “Le château des Albret à Labrit (Landes)”, in : Résidences aristocratiques, résidences du pouvoir entre Loire et Pyrénées, Xe-XVe siècles. Recherches archéologiques récentes, 1987-2002, Archéologie du Midi médiéval, 4, 337-363.
- Lassure, J.-M. (1995) : De la Tour-Salle au Castelnau : une étape du peuplement en Gascogne : les fouilles archéologiques médiévales de Corné à L’Isle-Bouzon (Gers), thèse de doctorat, Université Toulouse II.
- Lassure, J.-M. (1998) : La civilisation matérielle de la Gascogne au XIIe et XIIIe siècle. Le mobilier du site archéologique de Corné à l’Isle-Bouzon (Gers), thèse de Doctorat, Université Toulouse II.
- Lawler, D. F., Widga, C., Rubin, D. A., Reetz, J. A., Evans, R. H., Tangredi, B. P., Thomas, R. M, Martin, T. J., Hildebolt, C., Smith, K., Leib, D., Sackman, J. E., Avery, J. G. et Smith, G. K. (2016) : “Differential diagnosis of vertebral spinous process deviations in archaeological and modern domestic dogs”, Journal of Archaeological Science : Reports, 9, 54-63.
- Lemagnen, S. (2009) : “La tapisserie de Bayeux : une chronique des temps vikings ? Actes du colloque international de Bayeux 29 et 30 mars 2007”, Rouen.
- Lescure, S., Arnaud-Fassetta, G. (2015) : “Paléo-environnement et contrainte fluviale à l’Holocène récent sur les sites de Langoiran et d’Isle-Saint-Georges : bilan de quatre années de recherches géoarchéologiques dans la basse vallée de la Garonne”, Aquitania, 5, 43-64.
- Lescure, S., Arnaud-Fassetta, G., Le Coeur, C., Colin, A., Faravel, S., Mathé, V., Montenegro, M.-E. et Carozza, J. M. (2015) : “Évolution hydrogéomorphologique holocène de la Garonne maritime – Focus sur les sites de Langoiran et de l’Isle-Saint-Georges (Gironde, France)”, Géomorphologie : relief, processus, environnement, 21, 1, 57-72.
- Lignereux, Y. (2011) : “Nourritures carnées au Moyen-Âge – situation dans les castra entre Pyrénées et Languedoc (XIIIe-XVe s.)”, Bulletin du Centre d’Étude d’Histoire de la Médecine, 77, 27-54.
- Mallye, J.-B., Costamagno, S., Boudadi-Maligne, M., Prucca, A., Laroulandie, V., Thiebaut, C. et Mourre, V. (2012) : “Dhole (Cuon alpinus) as a bone accumulator and new taphonomic agent? The case of the Noisetier Cave (French Pyrenees)”, Journal of Taphonomy, 10, 3-4, 317-347.
- Martinaud, M. et Faravel, S., dir. (2007) : Le Castéra et le Palus de Langoiran (Gironde), rapport de prospection géophysique 2006, Service Régional de l’Archéologie d’Aquitaine, Bordeaux.
- Parkinson, J.A., Plummer, T. et Bose, R. (2014) : “A GIS-based approach to documenting large canid damage to bones”, Palaeogeography, Palaeoclimatology, Palaeoecology, 409, 57-71.
- Poplin, F. (1977) : “Problèmes d’ostéologie quantitative relatifs à l’étude de l’écologie des hommes fossiles”, Bulletin de l’Association Française pour l’Étude du Quaternaire, 47, 63-68.
- Poplin, F. (1979) : “L’origine du chien”, Ethnozootechnie, 25, 7-11.
- Prodéo, Fr., dir. (2007) : Pineuilh (33) ‘La Mothe’, RFO, Inrap Grand Sud-Ouest.
- Prucca, A. (2003) : “Caractérisation de l’impact des loups sur des ossements d’herbivores (cerfs de Virginie, orignaux, bisons) : étude des modifications infligées par les loups captifs et sauvages Nord-américains”, mémoire de DEA, Université de Provence, Aix-Marseille.
- Rambourg, P. (2012) : “Le menu du Moyen Âge au XXe siècle : témoin de l’histoire et de la gastronomie”, http://patrimoine.bm-dijon.fr/pleade
- Reitz, E. J. et Wing, E. S. (1999) : Zooarchaeology. Cambridge.
- Renou, S. (en préparation) : “Étude de la faune issue de la fouille du site de La Rochelle, ‘Rue des Augustins’ (Charente-Maritime, 17)”, in : Gissinger, B., La Rochelle, ‘Rue des Augustins’, Rapport de fouille archéologique préventive, Conseil général de la Charente-Maritime.
- Rodet-Belarbi, I. et Forest, V. (2009) : “Alimentation carnée du XIVe au XVIe siècle dans le sud de la France, d’après les sources archéozoologiques : la part des mammifères sauvages et des oiseaux”, in : F. Ravoire et A. Dietrich, éd., 2009, La cuisine et la table dans la France de la fin du Moyen Âge : contenus et contenants du XIVe au XVIe siècle – Actes du colloque organisé par l’UMR 5594, Dijon, l’Inrap, et le Centre d’étude et de recherche du patrimoine de Sens, Sens, 8-10 janvier 2004, 125-146.
- Yvinec, J.-H. (1996) : “Éléments de synthèse sur l’alimentation carnée durant le haut Moyen Âge dans le Douaisis”, in : Colardelle, M., éd., 1996, L’homme et la nature au Moyen Âge. Paléoenvironnement des sociétés occidentales, Paris, 30-35.
Notes
- Audoin-Rouzeau 1993 ; Clavel 2001 ; Durand et al. 1997 et Rodet-Belarbi & Forest 2009.
- Lignereux 2011 ; Rambourg 2012 et Renou en préparation.
- Beuchet & Clavel 2014 ; Borvon 2012 ; Rodet-Belarbi & Forest 2009 et Yvinec 1996.
- Beck 1984.
- Audouin 1986 et Bodson 1979.
- d’Anthenaise 2002 ; Baratay 2003 et Poplin 1979.
- Baratay 2003 ; Collectif BnF ; Dumolin 1935 ; Lemagnen 2009 et Renou et al. ce volume.
- Araguas & Barraud 2006 ; Bourgeois 2009 et Caillat & Laborie 1997.
- Faravel en préparation et BSR 2016.
- Faravel et al. en préparation.
- Araguas & Faravel 2006.
- Didierjean 1988 et Faravel 2014.
- Sous la direction de Sylvie Faravel.
- Faravel 2004 et Martinaud & Faravel 2007.
- Cf. notamment Faravel 2015.
- Faravel 2014.
- Cf. fouilles de Labrit (40), Laborie 2006 et de Pineuilh (33), Prodeo 2007.
- Buffière 2015.
- Lassure 1995.
- Bourgeois 2009.
- Renou et al. ce volume.
- Lescure & Arnaud-Fassetta 2015 et Lescure et al. 2015.
- Faravel 2015.
- Les restes appartenant à la phase VIb correspondant à la phase d’abandon et de mise en culture du site (129) et ceux recensés hors stratigraphie (63), ne sont pas considérés pour la suite de l’étude.
- Étude en cours et conduite par Brice Ephrem pour les restes de poissons.
- Hesse & Wapnish 1985 ; Klein & Cruz-Uribe 1984 et Reitz & Wing 1999.
- Brugal et al. 1994 et Poplin 1977.
- Modifié d’après Costamagno 1999 et Fosse 1994.
- Binford 1981 ; Blumenschine et al. 1996 ; Capaldo & Blumenschine 1994 et Fisher 1995.
- d’Errico & Villa 1997.
- Renou et al. ce volume.
- Faravel 2015.
- Harcourt 1974.
- Boudadi-Maligne 2010.
- Renou et al. ce volume.
- Goret et al. 2009.
- Lawler et al. 2016.
- Campmas & Beauval 2008 ; Fosse et al. 2011 et Parkinson et al. 2014.
- Campmas & Beauval 2008 ; Castel 2004 et Prucca 2003.
- Clavel 2001.
- Dominguez-Solera & Dominguez-Rodrigo 2009 et Greenfield 1988.
- Esteban-Nadal et al. 2010 ; Barja Núñez & Corona-M. 2007 ; Mallye et al. 2012.
- Mallye et al. 2012.
- Dominguez-Solera & Dominguez-Rodrigo 2009 et Greenfield 1988.
- Blancou 2000.
- “Perigòrd, meitat chen, meitat pòrc” (occitan, en français : moitié chien, moitié porc). Une expression souvent utilisée naguère pour caractériser les habitants du Périgord. Elle évoquerait la réputation d’habileté des Périgordins à localiser une truffe sans l’aide, ni d’un chien (=chen), ni (surtout) d’une truie (=pòrc) qui sont généralement utilisés pour cela.
