




Introduction et problématique
Que deviennent les amas de boucherie laissés par les groupes de chasseurs-cueilleurs après l’abandon de leur camp ? Si certains gisements exceptionnels permettent de reconstituer l’agencement des espaces de vie de groupes humains du passé et de pousser leur étude jusqu’à des questions d’ordre palethnologique1, la majorité des sites paléolithiques découverts livrent des ensembles osseux désorganisés du fait de processus post-dépositionnels variés2. De ces ensembles osseux, nous parvenons à déchiffrer les stratégies d’approvisionnement des espèces chassées, les modalités de transport des carcasses mais aussi les pratiques de boucherie opérées jusqu’aux choix de consommation (au sens le plus large du terme) usitées par les chasseurs-cueilleurs. Pourtant, les accumulations osseuses produites par les activités de boucherie des groupes humains du Paléolithique ont pu être la source de nourriture d’autres animaux. Si de nombreux travaux visant à mesurer l’impact des grands carnivores africains (hyène, lion) sur des restes osseux d’ongulés sont aujourd’hui disponibles3, un nombre plus limité concerne les carnivores évoluant hors de ce continent4. Le constat est encore plus alarmant en ce qui concerne les petits carnivores5.
Pour pallier ce manque de données dans le contexte de l’archéologie européenne, une expérimentation visant à mesurer le degré de perturbation que les loups peuvent provoquer sur un amas de boucherie a été conduite au parc “Les Loups du Gévaudan”.
Le site expérimental
“Les Loups du Gévaudan” est un parc animalier implanté sur la commune de Saint-Léger-de-Peyre en Lozère (Fig. 1). Dans une partie ouverte au public, le parc comporte plusieurs sous-espèces de loups de différentes régions du globe dont la Pologne (Canis lupus lupus, une meute), la Sibérie (C. l. albus, une meute), le Canada (une meute de Canis lupus arctos et une meute de C. l. occidentalis), la Mongolie (C. l. chanco, une meute). En marge du parc animalier se trouve un parc scientifique de 12 ha dans lequel évoluait, au moment de notre expérimentation, une cinquantaine de loups de Mongolie répartis en deux meutes. Les loups sont nourris tous les deux jours de morceaux de carcasses d’ongulés de différentes tailles (bœuf, mouton, porc). Occasionnellement des carcasses entières de chevreuil, mouton ou de cerf leur sont données.

Matériel et méthode
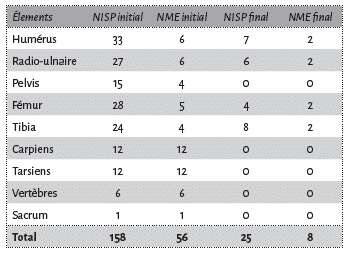
Cette expérience s’est déroulée au mois d’octobre 2010. Pour cette expérimentation, nous avons utilisé des os de mouton (NME initial = 56) issus de boucherie. Ces os ont été préparés le 24 octobre. La chair attenante à l’os et les tendons ont été enlevés à l’aide de couteaux en métal puis les os ont été fracturés avec un percuteur en pierre et une enclume pour en extraire la moelle. Ainsi avons-nous obtenu 158 fragments osseux (Tab. 1). Chaque fragment a été enregistré dans un tableur après avoir été numéroté par gravure mécanique.
Les os ont ensuite été déposés le 26 octobre tôt le matin dans l’enclos (Fig. 1) en neuf bandes, chaque os étant distant des autres de 30 cm. La position de chaque os a été enregistrée spatialement de manière à mesurer leur dispersion après l’intervention des loups.
Le lendemain, la cellule a été contrôlée et démontée. Le sol a été soigneusement inspecté dans un rayon de 20 m autour du dépôt initial. La position spatiale de chaque os a été relevée afin de calculer les distances de déplacements. Les os ont ensuite été bouillis à l’eau claire afin d’enlever les derniers lambeaux de chair et la graisse facilitant ainsi leur manipulation et l’observation des traces laissées par les charognards.
Les surfaces osseuses ont été minutieusement inspectées à la recherche de traces de carnivore à l’aide d’une loupe binoculaire (grossissement x40). La perte des éléments a été mesurée selon le pourcentage de changement (“Percentage change”)6 calculé de la manière suivante : (NME initial – NME final) / (NME initial) x 100. Plus le pourcentage est élevé, plus la perte osseuse est importante et moins la portion peut être considérée pour estimer l’abondance d’un élément.
Résultats
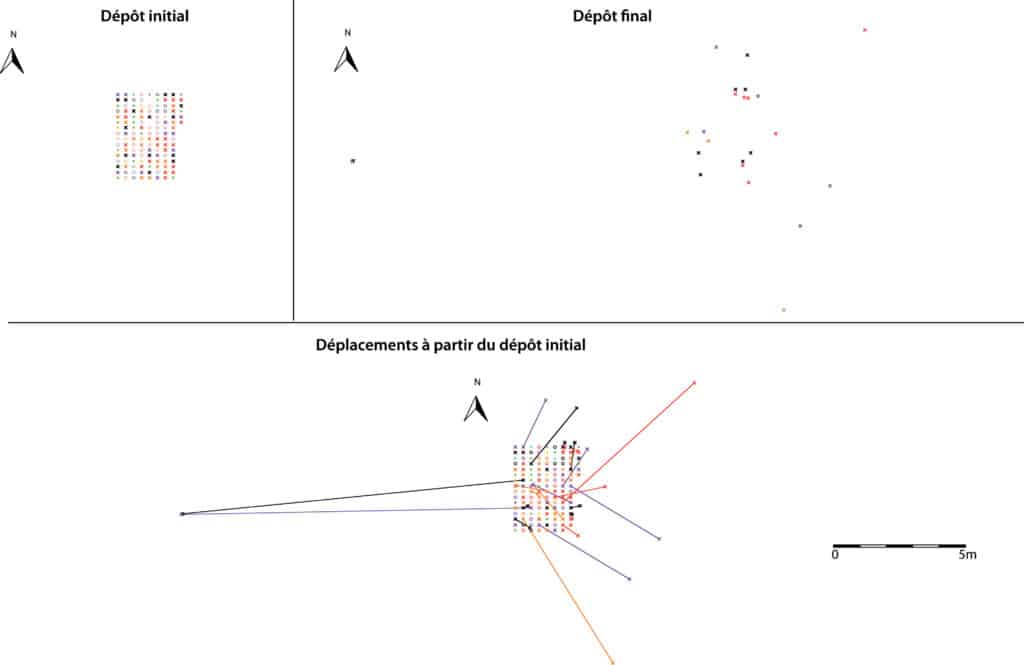
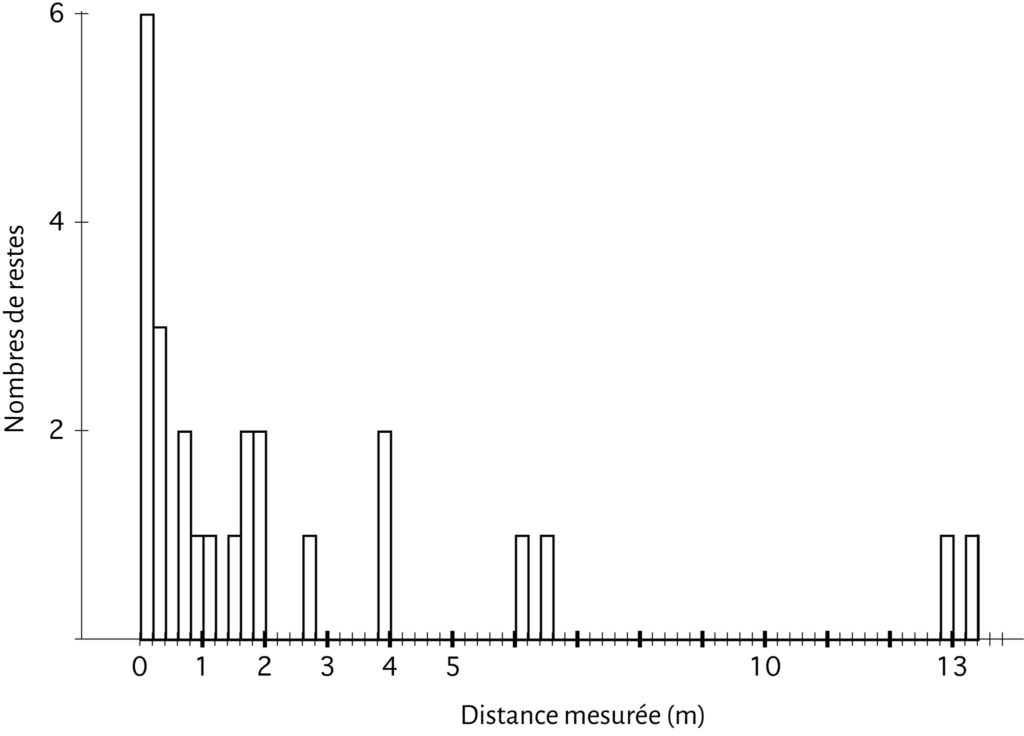
Des 158 os déposés, seuls 25 ont été retrouvés, correspondant à un nombre d’éléments final de 8. La perte osseuse est donc immense (84 % du NR, 85 % du NME) et l’amas osseux initialement déposé n’est plus apparent (Fig. 2). Si un quart des ossements recueillis n’ont été que peu déplacés (< 20 cm), les autres vestiges ont été récupérés à une distance importante de leur position initiale et parfois à plus de 10 m (Fig. 3, 4).

Tous les restes retrouvés sont des fragments de diaphyse tandis que toutes les parties articulaires des os longs mais aussi tous les éléments des ceintures et du squelette axial ainsi que les carpiens et les tarsiens ont disparu. Les pourcentages de perte calculés montrent que les valeurs les plus faibles sont toujours obtenues pour les portions de diaphyse moyenne et, dans une moindre mesure, pour les fragments proximaux et distaux de diaphyse (Fig. 5). Enfin, l’analyse des surfaces osseuses a révélé que seul un ossement parmi les 25 restes retrouvés, portait des traces de dents (Fig. 6). Il s’agit de la face latérale d’un fragment de diaphyse d’un tibia droit de mouton qui porte une série d’enfoncements sub-circulaires. Ce fragment a été découvert à une soixantaine de centimètres de sa position initiale.
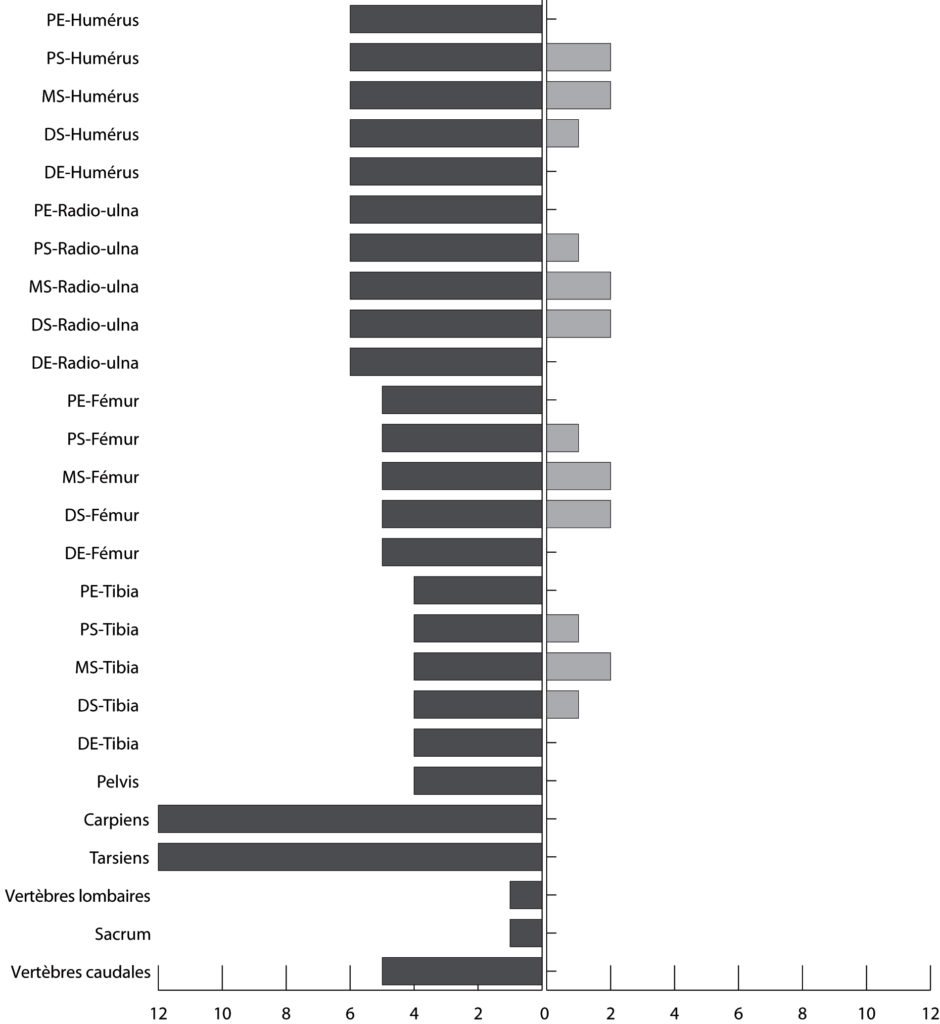
PE : extrémité articulaire proximale ; PS : diaphyse proximale ; MS : diaphyse moyenne ; DS : diaphyse distale ; DE : extrémité articulaire distale.

Discussion
Cette première expérimentation témoigne d’un impact non négligeable des loups sur l’agencement des amas de boucherie, soulevant de multiples questions quant à l’interprétation des ensembles osseux fossiles. En premier lieu, les déchets de boucherie utilisés dans cette expérience ont suscité un intérêt nutritif suffisant pour être prélevés par les loups. La consommation d’animaux morts autrement que par la chasse par le loup ou la consommation de cadavres humains a par ailleurs été notée plusieurs fois à travers le monde7 ; nos résultats n’ont donc rien de surprenant. Néanmoins, la taille de l’échantillon demeure faible par rapport à ce qui était laissé par les groupes de chasseurs-cueilleurs et l’est tout autant au regard du nombre de loups composant la meute de l’enclos. C’est un biais non négligeable qui pourrait être responsable de ce taux de perte si important.
L’amas originel a été complètement déstructuré en l’espace de seulement 24 heures ce qui est très rapide même si des résultats comparables ont été obtenus dans le cadre d’expériences conduites avec des hyènes8. Pour des questions purement pratiques et matérielles visant à mesurer rapidement la dispersion des pièces après le passage des canidés, les os de cette cellule ont été déposés en ligne et espacés les uns des autres. Cette configuration était donc très éloignée d’un amas abandonné après des opérations de boucherie. Les os étant très accessibles pour les prédateurs, les résultats pourraient être différents si les pièces osseuses avaient été déposées en tas. Dans cette expérimentation, nous n’avons pas utilisé de caméra de surveillance à détecteur de mouvement, ce qui aurait pu être utile afin de connaître non seulement le nombre de loups impliqués dans le chapardage des restes mais aussi pour évaluer la vitesse de dégradation de l’amas déposé. Ce dispositif permettrait en outre de mesurer le rôle des oiseaux charognards, tels que le vautour ou le grand corbeau, aussi présents autour du parc, dans la modification des cellules expérimentales.
Dans plusieurs cas expérimentaux, l’impact des chapardeurs a pu être observé et mesuré aux dépens des analystes ; ainsi dans le cadre d’une expérimentation visant à mesurer l’altération des os soumis aux conditions périglaciaires, des cellules osseuses déposées en haute altitude ont connu une perte de matériel de 50 % sur des os frais dès la première année d’expérimentation9. Les pertes occasionnées par des charognards sur des restes aviaires déposés dans le cadre d’une expérience conduite par Davis et Briggs10 sont elles aussi importantes, de l’ordre de 30 à 40 %.
Tout comme dans le cas des grands carnivores africains, les loups sont capables de modifier de manière significative la composition d’un ensemble osseux que cela soit en termes du nombre d’ossements ou de la représentation des différentes portions d’un os. Les taux de perte les plus faibles sont obtenus sur les portions de diaphyses moyennes. Dans notre cas cependant, la perte de matériels apparaît beaucoup plus conséquente ; le pourcentage de changement est toujours supérieur à 50 % alors qu’il est compris entre 0 et 82 % dans les précédentes expérimentations11. Le taux élevé de perte observé au Gévaudan pourrait s’expliquer par la quantité relativement faible d’ossements qui a été déposée et par la nature même des parties squelettiques qui sont représentées. Il est en effet probable que ce taux de perte aurait été différent si des parties moins riches en graisse avaient été déposées. Ce taux élevé pourrait aussi s’expliquer par la nature même du dépôt. Dans notre cas, les os étant dispersés plusieurs loups ont pu interagir simultanément sur la cellule. Si la cellule avait consisté en un amas d’os disposés sur un espace plus réduit, un ou peut-être deux loups auraient pu s’approprier cette cellule au détriment des autres membres de la meute et passer plus de temps à les mâchonner.
La fréquence des traces de dent dans cette expérimentation est relativement faible ce qui tendrait à montrer qu’il peut être difficile d’établir un lien entre le pourcentage de traces de carnivores et la perte osseuse générée par leur activité. Ceci souligne d’autant plus la difficulté d’identifier les actes de chapardage lors des analyses des séries fossiles. La multiplication des expériences permettra néanmoins de confirmer ces premières tendances. En effet, les vestiges avec un fort attrait nutritif étant très facilement accessibles pour les loups du fait de la façon dont a été constituée la cellule expérimentale (lignes d’ossements et non tas d’ossements), il n’est pas exclu que ce très faible pourcentage soit lié à cette configuration particulière. Ici encore dans le cas d’un amas en tas, un nombre plus réduit de loup (couple dominant ?) aurait eu accès à ces ressources et en s’installant près de l’amas aurait eu tout le loisir de mâchonner les ossements conduisant de fait à un taux plus élevé de traces.
Enfin l’expérimentation s’est centrée autour de restes osseux d’ongulé de taille modeste (mouton). Le tableau de chasse des chasseurs-cueilleurs étant très varié dans l’espace et dans le temps, gageons que d’autres expérimentations du même type puissent être entreprises sur les restes osseux d’ongulés de taille variée et des restes de petite faune.
Remerciements
Nous tenons à remercier Joseph Matera, directeur du parc “Les Loups du Gévaudan” de nous avoir donné accès aux enclos pour mener à bien nos expérimentations sur les loups. Nous remercions également toute l’équipe du Parc pour son aide avec une mention spéciale à Sylvain Macchi pour son immense aide et la transmission de ses connaissances sur le Loup. Lysianna Ledoux nous a apporté une aide non négligeable sur le terrain, qu’elle soit également remerciée. Nous souhaitons enfin remercier les deux rapporteurs de cet article qui ont contribué à améliorer la première version du manuscrit.
Bibliographie •••
- Arriaza, M. C., Dominguez-Rodrigo, M., Yravedra, J. et Baquedano, E. (2016) : “Lions as Bone Accumulators? Paleontological and Ecological Implications of a Modern Bone Assemblage from Olduvai Gorge”, PLoS ONE, 11, 5
- Binford, L. R. et Bertram, J. B. (1977) : “Bone Frequencies And Attritional Processes”, in : Binford, L. R., éd., For Theory Building in Archaeology. Essays on Faunal Remains, Aquatic Resources, Spatial Analysis, And Systemic Modeling, 77-153.
- Blumenschine, R. J. (1988) : “An experimental model of the timing of Hominid and carnivore influence on archaeological bone assemblages”, Journal of Archaeological Science, 15, 483-502.
- Capaldo, S. D. (1998) : “Simulating the formation of dual-patterned archaeofaunal assemblages with experimental control samples”, Journal of Archaeological Science, 25, 4, 311-330.
- Capitani, C., Bertelli, I., P., V., Scandura, M. et Apollonio, M. (2004) : “A comparative analysis of wolf (Canis lupus) diet in three different italian ecosystems”, Mammalian Biology, 69, 1, 1-10.
- Demski, M. (2015) : Kill or scavenge. The use of alternative food source in Scandinavian wolves, Master Thesis, Uppsala University.
- Dominguez-Rodrigo, M. (1999) : “Flesh availibility and bone modifications in carcasses consumed by lion : palaeocological relevance in hominid foraging patterns”, Palaeogeography, Palaeoclimatology, Palaeoecology, 149, 373-388.
- Liu, B. et Jiang, Z. (2003) : “Diet composition of wolves Canis lupus in the northeastern Qinghai-Tibet Plateau, China”, Acta Theriologica, 48, 2, 255-263.
- Marean, C. W. (1991) : “Measuring the Post-Depositional Destruction of Bone in archaeological Assemblages”, Journal of Archaeological Science, 18, 677-694.
- Marean, C. W. et Bertino, L. (1994) : “Intrasite spatial analysis of bone: subtracting the effect of secondary carnivore consumers”, American Antiquity, 59, 4, 748-768.
- Marean, C. W. et Spencer, L. M. (1991) : “Impact of Carnivore ravaging on zooarchaeological measures of element abundance”, American Antiquity, 56, 4, 645-658.
- Marean, C. W., Spencer, L. M., Blumenschine, R. J. et Capaldo, S. D. (1992) : “Captive Hyaena bone choice and destruction, the schlepp effect and Olduvai archaeofaunas”, Journal of Archaeological Science, 19, 101-121
- Mohammadi, A., Kaboli, M., Sazatornil, V. et López-Bao, J. V. (2019) : “Anthropogenic food ressource sustain wolves in conflict scenarios of Western Iran”, PLoS ONE, 16, 6, p. e0218345.
- Nascou, A. et Morin, E. (2014) : “Arctic wolf and spotted hyena gnawing damage on an experimental faunal assemblage”, Journal of taphonomy, 12, 1, 1-36.
- Nascou, A. L. (2012) : Variation in arctic wolf (Canis lupus arctos) and spotted hyena (Crocuta crocuta) gnawing damage on an experimental faunal assemblage. Master Thesis, Trent University.
- Thompson, J. C. et Lee-Gorishti, Y. (2007) : “Carnivore Bone Portion Choice and Surface Modification on Modern Experimental Boiled Bone Assemblage”, Journal of Taphonomy, 5, 3, 121-135.
Notes
- Leroi-Gourhan & Brézillon 1983 ; Audouze et al. 1981 ; Julien & Karlin 2014 ; Biard et al. 2018.
- Wood & Johnson 1978 ; Texier 2000.
- Blumenschine 1988 ; Marean 1991 ; Marean & Spencer 1991 ; Marean & Bertino 1994 ; Marean et al. 1992 ; Capaldo 1998 ; Dominguez-Rodrigo 1999 ; Thompson & Lee-Gorishti 2007 ; Nascou 2012 ; Nascou & Morin 2014 ; Arriaza et al. 2018.
- Binford & Bertram 1977 ; Nascou 2012 ; Nascou & Morin 2014.
- Leoni et al., ce volume.
- Marean & Spencer 1991 ; Marean et al. 1992.
- Liu & Jigang 2003 ; Capitani et al. 2004 ; Demski 2015 ; Mohammadi et al. 2019.
- Binford et al. 1988 ; Marean & Bertino 1994 ; Thompson & Lee-Gorishti 2007 ; Arriaza et al. 2018.
- Mallye et al. 2009.
- Davis & Briggs 1998.
- Mallye et al. 2009.
