




Introduction
Au cours de la Protohistoire, l’animal est capital puisqu’il fournit force de travail, nourriture et matières premières. L’étudier revient donc à appréhender les besoins des populations anciennes, mais aussi leurs possibilités, directement liées à leurs connaissances et à leur environnement. Les pratiques d’élevage et les activités cynégétiques sont donc susceptibles d’évoluer dans le temps, de varier dans l’espace, ou encore en fonction de l’aisance économique ou des exigences d’une population. Or, si des études ont bien été menées au fil des découvertes ces quarante dernières années, la réalisation d’une synthèse sur les faunes du Premier âge du Fer était nécessaire pour mieux appréhender ces activités et, plus globalement, les rôles et importances des animaux au sein de ces sociétés hallstattiennes.
Dans ce cadre, une zone géographique relativement vaste, qui correspond au nord-est de la France, a été considérée. Il s’agit, somme toute, de la zone de diffusion de la culture hallstattienne sur le territoire français actuel (Fig. 1). Plusieurs régions sont donc concernées : la Lorraine, l’Alsace, la Bourgogne, la Franche-Comté, la Champagne-Ardenne et l’est de la région Centre. Grâce à différents axes de circulation, aussi bien terrestres que fluviaux, cette zone constitue une véritable aire de transit de biens et de personnes au cours du Premier âge du Fer, aussi bien du sud au nord de l’Europe que d’est en ouest.

(d’après B. Chaume et W. Reinhard, in Chaume & Mordant 2001).
L’obtention d’une vision d’ensemble de la place de l’animal dans la société hallstattienne nécessitait, en outre, de considérer deux grands types d’habitats que sont les établissements ruraux ouverts d’une part, et les sites aux statuts plus particuliers d’autre part. Nommés “résidences aristocratiques”, ces derniers sont des habitats fortifiés, situés à proximité d’axes de circulation, sur des hauteurs et occupés à des périodes relativement longues.
Ce travail, mené dans le cadre d’un doctorat, se propose donc d’établir un premier bilan des données archéozoologiques du Premier âge du Fer dans le nord-est de la France, dont les premiers résultats sont présentés ici.
L’étude d’une période de près de quatre siècles et la prise en compte d’une zone géographique étendue ont nécessité l’établissement d’un premier inventaire des lots de faunes disponibles dans ces régions, étudiés ou non, et associés à des contextes d’habitat. Si cet inventaire n’est pas exhaustif à l’heure actuelle – en ce qui concerne les lots non étudiés ou récemment analysés notamment – il tend à montrer l’ampleur de la documentation archéozoologique d’ores et déjà disponible puisqu’il est riche de cent-vingt-cinq sites d’habitat qui s’échelonnent entre le Hallstatt C et La Tène A (Fig. 2, en haut). La plupart des lots provient d’occupations domestiques rurales de petites envergures mais l’on compte également sept habitats de hauteur fortifiés ainsi qu’un centre proto-urbain, Bourges “Avaricum”.
Les habitats ruraux sont nombreux, mais livrent souvent moins de cinq cent restes, ce qui a des conséquences sur la représentativité des lots. Ces vestiges sont, en outre, répartis dans plusieurs structures qui ne sont pas toujours synchrones. Les données archéozoologiques acquises à partir de ces ensembles sont donc susceptibles de présenter des biais, d’autant plus qu’à l’inverse, les habitats de hauteur livrent parfois plus de dix mille restes osseux qui peuvent être rattachés à une même occupation. Ces écarts d’effectifs résultent de divers facteurs, parmi lesquels la durée et l’ampleur des fouilles archéologiques ont leur importance. En effet, les habitats de hauteur ont bénéficié de campagnes de fouilles sur de longues durées, parfois plus de dix ans, tandis que les habitats ruraux sont découverts et la plupart du temps traités dans le cadre d’opérations préventives. Par ailleurs, les habitats de plaine n’ont pas suscité le même engouement que les habitats fortifiés au cours du XXe s. et ne sont finalement assez bien connus que depuis la systématisation des opérations de diagnostic et de fouilles préventives.
Il faut également considérer les durées d’occupation, les habitats de hauteur étant généralement occupés sur de longues périodes, à l’inverse des occupations rurales qui ont tendance à se déplacer au fil du temps.
D’autre part, et c’est sans doute là un facteur déterminant, les habitats de hauteur fortifiés sont susceptibles d’accueillir un nombre relativement important d’occupants, contrairement aux occupations rurales qui ne comptent généralement que quelques bâtiments. Enfin, les modes de gestion des déchets, qui conditionnent la conservation et la préservation des restes osseux, varient entre les habitats de hauteur où ils sont soit entassés en bordure des remparts et ainsi maintenus et préservés au fil du temps, soit rejetés dans des structures fossoyées, et les occupations rurales où la plupart d’entre eux sont probablement simplement déposés sur les tas de fumier ou à même le sol.
Les cent-vingt-cinq ensembles référencés sont donc marqués par des différences d’effectifs, auxquelles il faut ajouter des différences de conservation et de représentativité. Tous ces éléments ont conduit à ne considérer que les sites fournissant un nombre minimum de 200 restes déterminés. Outre ce critère quantitatif, l’ampleur de la documentation archéozoologique alliée aux délais impartis à cette étude ont amené à sélectionner les ensembles sur la base de critères chronologiques et contextuels. Au sujet, tout d’abord, des sites dont les restes animaux n’avaient pas été étudiés, la priorité a été donnée aux ensembles bien datés issus des habitats de hauteur : le “Camp de la Roche ” à Bourguignon-les-Morey, le “Camp du Château” à Salins-les-Bains, le “Britzgyberg” à Illfurth, le “Camp d’Affrique” à Messein et le “Kastelberg” à Koestlach-Moernach. Un ensemble provenant d’un habitat rural a également été étudié puisqu’il avait livré de nombreux restes issus de structures bien datées (“In der Klamm/Schwobenfeld” à Entzheim-Geispolsheim, Bas-Rhin). En ce qui concerne les ensembles étudiés par d’autres auteurs, n’ont été retenus que ceux qui étaient suffisamment documentés et qui comportaient plus de deux cents restes déterminés.
Outre les critères chronologiques, contextuels et quantitatifs, des choix ont également été opérés sur la base des méthodes d’études archéozoologiques employées par d’autres spécialistes. En effet, dans certaines études anciennes, les effectifs totaux ne sont pas toujours renseignés, de même que les masses des restes, leur contexte, ou encore leur chronologie lorsque plusieurs occupations se succèdent sur un même site.
Finalement, quinze grands ensembles ont été pris en compte (Fig. 2, en bas), dont sept habitats de hauteur fortifiés qui ont livré de grandes quantités de restes animaux et qui ouvrent ainsi à des études individuelles et comparatives. Un centre proto-urbain, le promontoire de Bourges “Avaricum” a également été intégré, ainsi que sept habitats ruraux. Le corpus rassemble 159401 fragments osseux, dont 149457 issus d’habitats “privilégiés” et 9944 d’occupations de moindre envergure. Nous partons ainsi d’un corpus conséquent qui, s’il n’atteint pas l’exhaustivité géographique, permet d’aborder de nombreux points de manière représentative.
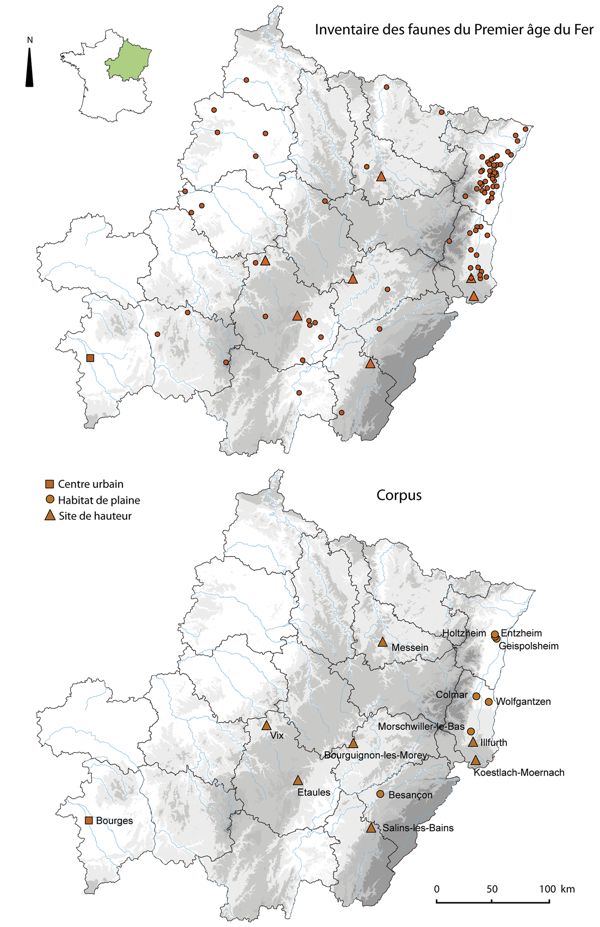
étudiés ou non (en haut) et distribution géographique des habitats du corpus (en bas).
Du point de vue de la chronologie, notons qu’avec trente ensembles ce corpus recouvre l’intégralité du Premier âge du Fer, soit une période de quatre siècles (800-400 a.C.). Aux écarts d’effectifs entre les habitats déjà énoncés, il faut ajouter des écarts d’effectifs entre les différentes phases. Commençons d’abord par préciser que si la plupart des contextes sont datés avec précision, certains – anciennement fouillés notamment – sont attribués à de larges plages chronologiques, comme le “Camp du Châtelet” à Étaules, dont l’étude de faune correspond à l’occupation du site au Hallstatt C et au Hallstatt D sans plus de précision. Si ces contextes à chronologie peu détaillée ont été pris en compte, c’est qu’ils ont l’avantage de fournir des quantités de restes animaux représentatives. En ce qui concerne les habitats de hauteur, tous à l’exception du “Kastelberg” sont occupés au cours du Hallstatt D2 et D3, ce qui offre de nombreuses possibilités de comparaisons. Le promontoire de Bourges “Avaricum” et cinq habitats de plaine sont, par ailleurs, également occupés au cours de cette phase. Au final, si l’ensemble de la période est représenté par des habitats de hauteur et des occupations de plaine, c’est au Hallstatt final (Hallstatt D2-D3) que se rapporte près de la moitié de la documentation, aussi bien en restes animaux (49 % des restes déterminés) qu’en ensembles chronologiques avec quinze occupations distinctes (Fig. 3).

En somme, si le corpus se compose de quinze grands ensembles répartis de manière relativement inégale au sein de la zone d’étude, il n’en est pas moins que la proximité géographique des habitats de hauteurs, ou encore de sites de hauteur et d’occupations rurales contemporaines, sont autant d’éléments qui offrent des possibilités de comparaisons et l’accès à une vision suffisamment précise de l’emploi des animaux, de l’élevage, de la consommation de viande ou encore des activités cynégétiques durant cette période.
Apports des fréquences des espèces et des âges d’abattage à l’étude de l’élevage
Afin de poser les bases de cette étude, notons que 98 % des restes osseux déterminés sont issus d’animaux domestiques. L’élevage prédomine donc très nettement dans l’acquisition des ressources carnées et des matières premières. La chasse, quant à elle, est pratiquée partout, mais fournit très peu de ressources. La faune sauvage est assez variée puisque douze espèces de mammifères sauvages1 et plusieurs espèces aviaires sont attestées2 (de manière erratique néanmoins). Cette diversité d’espèces suggère que les activités cynégétiques pouvaient avoir d’autres finalités que la seule recherche de ressources carnées, telles que l’élimination d’animaux susceptibles de porter atteinte aux troupeaux ou aux cultures, la récupération des fourrures et autres matières premières, ou encore la pratique d’un loisir sportif ou d’un entrainement guerrier. Il est d’ailleurs intéressant de constater que les activités cynégétiques sont mieux représentées dans les habitats ruraux où les restes osseux qui leur sont associés ne comptent jamais pour moins de 1 % des restes déterminés et atteignent même 32 % à Holtzheim (Bas-Rhin). Dans les habitats de hauteur et à Bourges “Avaricum”, les mammifères sauvages représentent moins de 1 % des restes déterminés dans la plupart des cas et, quoi qu’il en soit, ne dépassent jamais 4 %.
L’élevage est donc une activité majeure au Premier âge du Fer, dans tous les habitats étudiés. Nous nous concentrerons ici sur les problématiques qui lui sont liées et plus particulièrement sur les contextes privilégiés que sont les habitats de hauteur et Bourges “Avaricum”. Les données inhérentes aux habitats ruraux seront, quant à elles, ponctuellement présentées afin de distinguer ces deux types d’ensembles.
D’un point de vue général, les animaux domestiques sont susceptibles de fournir un riche panel de produits et de services. Il existe ainsi diverses formes d’élevages, divers types d’exploitations, qui traduisent, non seulement les possibilités et besoins des populations humaines, en lien avec l’environnement, les activités agricoles ou encore la démographie, mais aussi leurs goûts et préférences. Cerner les objectifs de l’élevage consiste donc à aborder les choix effectués par les populations anciennes en matière d’espèces élevées, d’animaux sélectionnés pour la reproduction, la production ou le travail, ou encore ceux qui sont destinés à l’alimentation carnée. Cela revient donc à analyser les fréquences des espèces, à évaluer les âges d’abattage des animaux et à aborder les distributions sexuelles au sein des troupeaux.
Dans le cas des habitats de hauteur, la première caractéristique à prendre en compte est leur hétérogénéité. Puisque ces habitats se différencient par leur superficie, leur organisation interne et, probablement aussi, leur démographie, les variations des compositions et des modes de gestion des troupeaux sont susceptibles d’en témoigner. La seconde interrogation majeure concerne le statut des populations des habitats privilégiés. Cet aspect peut être abordé par les divers mobiliers archéologiques (céramique, métallique, parures, armement), mais peut également être entrevu à partir des déchets de consommation carnée. En effet, une proportion élevée d’animaux jeunes témoigne d’un certain affranchissement des contraintes économiques, puisqu’un animal abattu avant l’âge adulte fournit une viande de qualité, mais en plus petite quantité et qu’il n’est à l’origine d’aucun produit secondaire ni d’aucune force de travail de son vivant. Enfin, les modalités d’approvisionnement des habitats de hauteur doivent être étudiées, puisqu’ils se situent au sommet de reliefs dont les superficies sont relativement restreintes, les rendant alors peu propices à une activité d’élevage.
Les fréquences des espèces domestiques dans les habitats
Nous ne considérons ici que les espèces “phares” de l’élevage que sont le bœuf, le porc, le mouton et la chèvre (ces deux dernières espèces étant regroupées sous le terme de caprinés de par la difficulté de distinguer leurs os). Les masses des restes n’étant pas toujours renseignées3, seules les fréquences en nombres ont été prises en compte dans les analyses globales. Quoi qu’il en soit, à l’exception des ensembles du “Mont Lassois” à Vix et du “Collège Littré” à Bourges “Avaricum” où le porc prédomine, c’est le bœuf qui livre la masse la plus élevée dans tous les habitats (à raison de 38 à 74 % de la masse totale des mammifères domestiques).
Une première analyse, menée sur toutes les occupations sans distinction chronologique (Fig. 4A), a révélé la distribution relativement homogène, et en cela remarquable, des occupations de Bourges au sein desquelles le porc est toujours mieux représenté. Or, son élevage ne nécessite pas de grands espaces et son alimentation peut consister en des restes de repas. Il n’est donc pas inhabituel de le trouver dans des proportions importantes dans des contextes à forte démographie, comme des centres urbains, notamment au Second âge du Fer ou encore à la période gallo-romaine (Foucras 2011 ; Lepetz 1996).

du bœuf, du porc et des caprinés dans les habitats.
Dans les habitats de hauteur, les fréquences des espèces sont relativement homogènes. Cependant, le secteur du grand bâtiment du mont Lassois élargit considérablement le domaine de variation par une prédominance très marquée du porc (67 %). Mais parce ce que l’implantation du grand bâtiment absidial hallstattien a perforé un niveau d’occupation du Bronze final IIIb, la datation des restes fauniques piégés dans les fossés et trous de poteaux est incertaine. Si l’on considère donc tous les ensembles liés aux habitats de hauteur, à l’exception de ce dernier, cette catégorie apparaît plus homogène encore (Fig. 4B). Le porc et le bœuf y sont souvent les plus fréquents, dans des proportions qui varient cependant légèrement.
Enfin, la diversité des ensembles correspondants aux habitats de plaine est intéressante, d’autant plus qu’ils se situent tous (à l’exception d’un4) dans un environnement similaire : la plaine d’Alsace. Si l’on se réfère aux résultats obtenus pour d’autres périodes en Gaule (Fig. 4C), ces variations de fréquences pourraient correspondre, de manière schématique, à différents types d’exploitations rurales : des habitats groupés, des fermes autarciques, ou encore des exploitations plus spécialisées.
Quoi qu’il en soit, la diversité observée dans les trois grandes catégories d’habitats peut être le fait : de variations chronologiques, traduire une adaptation de la consommation carnée à des besoins voire des contraintes économiques et/ou environnementales, des choix culturels ou encore les différences de statuts. Cette diversité peut également suggérer un fonctionnement en réseau de certains de ces habitats.
Le cas de Bourges “Avaricum”
Les quatre ensembles associés à ce vaste habitat groupé (“Saint-Martin-des-Champs”, “Collège Littré”, “Hôtel-Dieu” et “Rue de Sarrebourg”) ont été analysés indépendamment puisque les fréquences du porc ont d’ores et déjà révélé sa spécificité. Il ne semble pas exister de réelle évolution sur cette période relativement courte du Hallstatt D2 à La Tène A. Nous n’observons pas non plus de variations entre les zones d’habitat dense et les faubourgs. Cependant, les fréquences de caprinés et de chien sont plus importantes à “Saint-Martin-des-Champs” que dans les autres ensembles. Or, celui-ci est le seul qui ait livré des témoignages d’artisanat sur matières animales. Ces variations sont donc susceptibles d’être liées à la nature de l’occupation d’une zone qui n’est pas exclusivement dédiée à l’habitat. Par ailleurs, on observe des différences entre les deux ensembles de l’habitat dense du plateau que sont “Collège Littré” et “Hôtel-Dieu”, le porc étant moins fréquent dans le second. Cela pourrait refléter des écarts de statut entre ces deux ensembles, que les analyses des âges d’abattage auront tendance à démontrer, nous le verrons.
Les habitats de hauteur
En ce qui concerne les habitats de hauteur, les analyses de fréquences des mammifères domestiques selon la chronologie (du Hallstatt D1 à La Tène A) n’ont pas non plus révélé de dynamique évolutive commune. En revanche, le traitement des données dans un sériographe révèle des spécificités propres aux habitats eux-mêmes (Fig. 5). Sur ce type de graphique, la colonne de gauche permet de constater les différences d’effectifs globaux pour chaque ensemble (NRt), les barres grises correspondent aux effectifs par espèces dans chaque ensemble (en %NR), les barres noires expriment l’Écart Positif au Pourcentage Moyen (EPPM), autrement dit l’écart des fréquences par rapport à la moyenne.
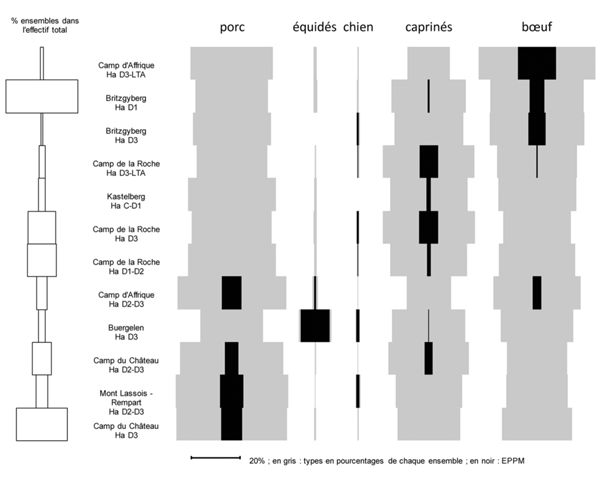
Les données sont sériées par le logiciel “ Sériographe EPPM 0.2” (Desachy 2004).
Ces graphiques permettent de distinguer trois grands groupes selon l’espèce dominante, soit le porc (“Camp du Château”, rempart du “mont Lassois” et première phase d’occupation du “Camp d’Affrique”), les caprinés (“Kastelberg” et “Camp de la Roche”) ou le bœuf (“Britzgyberg” et la seconde phase d’occupation du “Camp d’Affrique”). Il est intéressant de constater que le “Buergelen” se distingue nettement par un taux d’équidés élevé (13 %). Or, il est considéré comme le faubourg du “Britzgyberg” au Hallstatt D3 et sur ce site, c’est le bœuf qui prédomine (40 %) et les équidés sont absents. Ces éléments laissent envisager une certaine complémentarité entre ces deux occupations.
La chronologie n’étant pas en cause dans les variations de fréquences des mammifères domestiques, nous pouvons supposer que d’autres critères entrent en jeu, comme l’environnement naturel des sites et notamment l’altitude et la superficie des plateaux. En effet, l’élevage du bœuf peut s’avérer délicat en altitude puisqu’il nécessite de vastes herbages, de grandes quantités d’eau, mais surtout un apport fourrager ou céréalier les mois d’hiver, donc l’espace nécessaire à la culture de plantes fourragères ou une possibilité d’approvisionnement. À l’inverse, les moutons et chèvres s’adaptent mieux puisque leur alimentation est plus variée et qu’ils se contentent très bien des herbages pentus. Enfin, l’élevage porcin ne nécessite pas de grands espaces et leur régime alimentaire diversifié peut être contenté par les restes de repas. Or, on observe des variations qui correspondent assez bien aux contraintes et possibilités inhérentes à l’élevage de ces trois espèces à partir de 330 m (Fig. 6). Le bœuf est bien représenté entre 330 et 529 m, puis c’est le porc qui est prépondérant dans la catégorie suivante et enfin les caprinés à partir de 630 m. Cependant, le bœuf est également bien représenté dans la dernière catégorie. L’altitude à elle seule ne peut donc pas expliquer toutes les variations de fréquences observées.

Les données sont sériées par le logiciel “ Sériographe EPPM 0.2” (Desachy 2004).
Les superficies des plateaux, en revanche, pourraient être plus déterminantes, comme semblent l’indiquer plusieurs cas. En premier lieu le “Camp du Château” où le porc est l’espèce qui prédomine alors qu’il s’agit du plus petit habitat de hauteur connu à ce jour (0,75 ha) et que la topographie du plateau sur lequel il est installé est irrégulière et escarpée (Fig. 7). Le porc prédomine également au “mont Lassois”, où la table sommitale est relativement petite (5 ha).
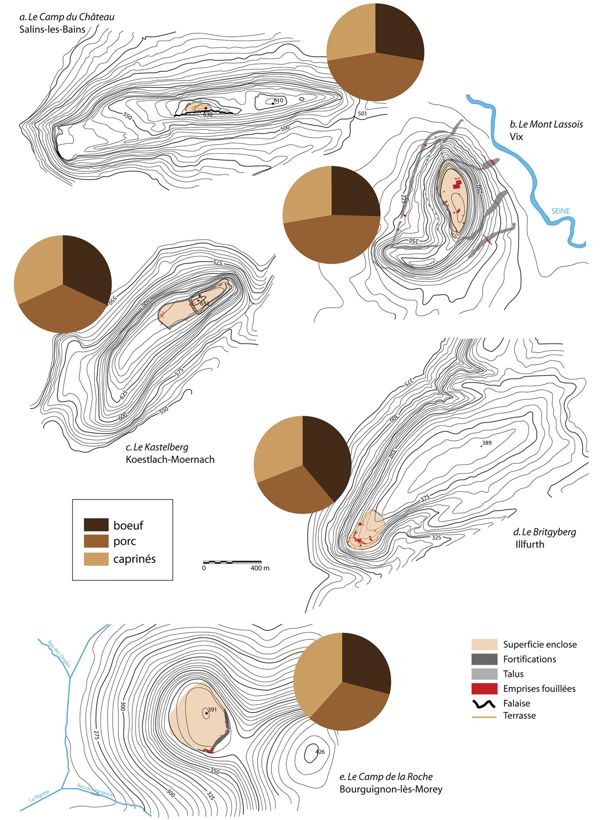
A. “Camp du Château”, Salins-les-Bains ;
B. “Mont Lassois”, Vix ;
C. “Kastelberg”, Koestlach-Moernach.
D. “Britzgyberg”, Illfurth ;
E. “Camp de la Roche”, Bourguignon-les-Morey.
Les variations de fréquences entre le “Britzgyberg” et le “Kastelberg” nous orientent également vers cette hypothèse. Ces deux sites sont géographiquement très proches, de superficies équivalentes et tous deux occupés au cours du Hallstatt D1, mais au “Britzgyberg”, le bœuf domine nettement (38 %), tandis qu’au “Kastelberg” ce sont les porcs qui sont les plus fréquents (36 %). Or, le plateau sur lequel est implanté le “Britzgyberg” est vaste et fait partie d’une chaîne de plateaux reliés entre eux par de faibles dénivellations, tandis que le “Kastelberg” est un relief isolé d’une vingtaine d’hectares et dont la table sommitale est pentue, ce qui limite les possibilités d’un élevage bovin in situ.
Enfin, au “Camp de la Roche”, qui compte parmi les plus vastes habitats de hauteur (17 ha), ce sont les caprinés qui dominent et non le bœuf. Ce relief est ceint de collines aux pentes relativement raides qui ne sont guère appropriées à l’élevage bovin mais qui pourraient convenir à des cheptels ovins et, plus sûrement encore, caprins. Ainsi, l’altitude des habitats, les superficies des plateaux, ainsi que leur topographie sont autant d’éléments qui influent sur les fréquences des espèces dans les habitats de hauteur. Mais ces facteurs n’expliquent pas toutes les variations qui sont également susceptibles de résulter de différences économiques ou démographiques. Celles-ci se reflèteront alors dans les gestions des cheptels et les qualités des viandes consommées.
Gestion des troupeaux et utilisations des animaux
L’élevage de mammifères domestiques, dont la finalité la plus évidente est l’obtention de ressources carnées destinées à l’alimentation humaine, est également à l’origine d’autres productions et de divers services. Parmi les plus développés, citons les productions laitières qui sont principalement le fait des bovins et des caprinés, la production lainière des ovins, les travaux de traction et de transport fournis par les bœufs et les équidés, le travail de protection ou de garde des cheptels des chiens et, enfin, la pérennité des troupeaux assurée par les reproducteurs. Aborder ces diverses productions, qu’elles soient matérielles ou non, nécessite de connaître les âges auxquels sont abattus les animaux. En effet, les animaux jeunes donnent une viande de qualité, mais seules les femelles adultes fournissent du lait et seuls les animaux dressés et entraînés sont aptes à tirer des charges.
Les données concernant les âges d’abattage des animaux d’élevage sont ici, pour l’essentiel, issues de l’observation du stade d’épiphysation des os (processus de soudure des différentes parties d’un même os au cours de la croissance). Ces estimations sont approximatives et donnent un âge théorique selon des classes dont les amplitudes, calées sur les âges d’épiphysation des os, varient selon les espèces.
L’élevage bovin, une production mixte
Dans le cas du bœuf, les estimations d’âges ont été regroupées en quatre catégories : de la naissance à 20 mois (veaux), les jeunes de 20 à 44 mois, les adultes de 44 mois à 8 ans et, enfin, les individus réformés de plus 8 ans (Fig. 8). Ces classes ont l’avantage de correspondre à diverses phases d’utilisations des animaux. En effet, la maturité pondérale du bœuf étant atteinte aux alentours de trois ans, il n’est pas nécessaire de conserver les animaux au-delà si l’élevage est uniquement destiné à la production de viande. Lorsque les animaux sont abattus avant d’avoir atteint cet âge, ils correspondent alors à des animaux de boucherie, dont la viande est tendre. Si des bêtes plus âgées sont conservées, cela indique une production laitière ou l’usage de la force de traction. Dans le cas présent, les données épiphysaires n’ont été prises en compte que lorsqu’elles étaient représentatives, ce qui limite l’étude à deux ensembles de Bourges “Avaricum”5 et six ensembles liés à des habitats de hauteur6. Par ailleurs, l’imprécision des méthodes d’estimation des âges d’abattage a conduit à placer la limite de maturité pondérale vers quatre ans (44 mois), on l’a vu, sans que cela ne perturbe outre mesure nos interprétations.
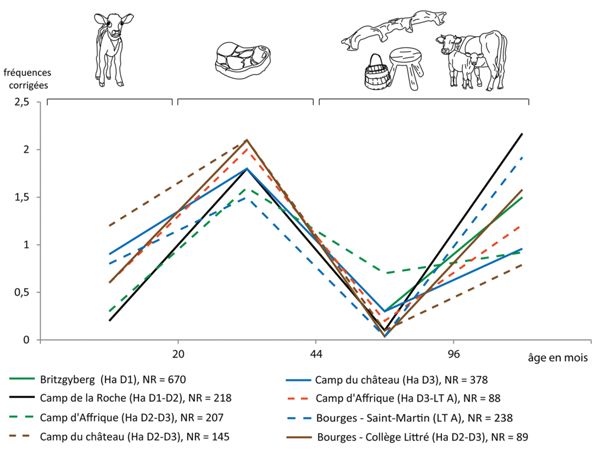
Les gestions des troupeaux bovins dans les habitats de hauteur et à Bourges sont similaires et traduisent une sélection d’animaux jeunes dans le cadre d’une production carnée de qualité puisque les animaux sont abattus avant leur maturité pondérale (entre 20 et 44 mois). Le lait et la force de traction sont néanmoins recherchés, c’est ce qu’attestent les fréquences des animaux abattus après réforme (plus de 8 ans). À Bourges, nous avons précédemment constaté des variations de fréquences des mammifères domestiques entre les ensembles du “Collège Littré” et de l’“Hôtel-Dieu”, tous deux associés à l’occupation dense du plateau. Or, dans le premier, la majorité des animaux est abattue jeune, alors qu’à “l’Hôtel-Dieu”, “la plupart des bovins a été abattue une fois adulte” (Augier et al. 2007). Ainsi, la zone du “Collège Littré” pourrait effectivement avoir eu un statut différent, plus privilégié peut-être. Dans certains cas, les estimations dentaires témoignent de sélections assez drastiques d’animaux très jeunes et de la rareté des bêtes de réforme. Ces éléments pourraient témoigner en faveur d’une importation d’animaux plutôt que d’un élevage in situ. C’est notamment vrai au “Camp du Château” au cours de sa seconde phase d’occupation (Hallstatt D3).
Dans les autres habitats de hauteur, les données sont trop peu nombreuses ou imprécises pour être intégrées à cette analyse. Notons tout de même qu’au “Camp de Châtelet”, la plupart des animaux est, a priori, abattue après avoir atteint la maturité pondérale, avec deux animaux de trois ans, trente-sept adultes et six individus âgés ou très âgés7 (Poulain, 2003). Les jeunes sont tout de même présents, avec notamment deux sujets de trois et quatre mois, deux autres de dix mois, cinq de vingt mois. Au “mont Lassois”, dans le secteur du rempart, “la majorité des restes provient de jeunes individus” (Huguet 2011b), traduisant une recherche de viande de qualité. Enfin, à Bourges “Hôtel-Dieu”, les données relatives aux stades d’épiphysation révèlent qu’aucun animal n’est âgé de moins de vingt mois et que 60 % d’entre eux ont été abattus après avoir atteint leur maximum de rendement carné (Augier et al. 2007). Or, les âges d’abattage des animaux au “Collège Littré”, qui fait également partie de l’agglomération, donnent des résultats inverses puisque la majorité des animaux est abattue avant trois ans. Ainsi, comme l’analyse des fréquences des mammifères domestiques le faisait déjà suspecter, la zone du “Collège Littré” semble avoir accueilli une population plus aisée que celle de l’“Hôtel-Dieu”.
Les données relatives aux âges d’abattage dans les habitats ouverts sont pour le moins lacunaires, lorsqu’elles ne sont pas absentes. Cela s’explique notamment par la taille réduite des ensembles. S’il n’est pas possible d’envisager les comparaisons avec les habitats de hauteur, ni même d’observer de grandes tendances parmi les habitats ouverts, nous pouvons mentionner qu’au “Simlisberg” (Hallstatt C), la plupart des ossements correspond à des animaux de plus de sept ans et les dents isolées proviennent d’adultes. Bien que quelques ossements puissent être attribués à de jeunes veaux, “l’impression d’ensemble est celle d’une population plutôt âgée” (Méniel 1991). De la même manière, à Colmar (Hallstatt D1), toutes les classes d’âges sont représentées, avec cependant davantage d’adultes (sept individus sur dix), dont deux âgés et un très âgé (Poulain 1990). Enfin, à Geispolsheim (Hallstatt D2-D3), 78 % des restes permettant une estimation des âges correspondent à des bêtes adultes, les très jeunes étant en revanche peu représentés (4 %) (Méniel 1990). Si les estimations sont trop peu nombreuses pour l’affirmer, nous pouvons remarquer que dans ces trois habitats ouverts, les animaux adultes et âgés sont toujours mieux représentés que les jeunes. En conséquence, et conformément à ce que les contextes et mobiliers archéologiques nous indiquent, l’économie dans les habitats ruraux paraît moins indépendante des contraintes de production et la viande bovine y semble de moindre qualité.
L’analyse de la gestion des cheptels bovins révèle donc des différences selon la nature des habitats. Bien que les données soient relativement ténues dans le cas des habitats ouverts, elles donnent l’image d’une production de bêtes de boucherie et, à plus forte raison, de vaches laitières et de bêtes de somme. La qualité de la viande bovine consommée y semble assez moyenne puisque très peu d’animaux sont abattus avant la maturité pondérale. Dans les habitats de hauteur et à Bourges, les situations traduisent le plus souvent une sélection d’animaux jeunes dans le cadre d’une production carnée de qualité. Il est intéressant de constater ici les différences de gestion entre les trois ensembles proto-urbains de Bourges “Avaricum”, “Collège Littré”, “Saint-Martin-des-Champs” et “Hôtel-Dieu”. Dans le premier, qui correspond à une zone d’habitat plutôt aisée, la viande bovine consommée est de bonne qualité, bien que les animaux de réforme soient présents. Les deux autres ensembles correspondent à une zone domestique et artisanale (“Saint-Martin-des-Champs”) et à une zone d’habitat (“Hôtel-Dieu”). Les animaux réformés y sont plus courants que les animaux jeunes, donnant ainsi l’image d’une consommation de viande de qualité moyenne et, donc, d’une aisance économique moindre.
Il se pourrait, par ailleurs, que les sélections d’animaux très jeunes à jeunes et la rareté des bêtes de réforme puissent traduire une importation d’animaux plutôt qu’un élevage in situ dans certains habitats de hauteur, on l’a vu. C’est notamment vrai au “Camp du Château” au cours de sa seconde phase d’occupation (Hallstatt D3), puisque la représentation de jeunes veaux y est particulièrement importante. Cependant, la présence d’animaux de tous âges ne permet que de le suspecter.
Le porc, animal de boucherie
Le porc occupe une place majeure dans l’alimentation des populations hallstattiennes. Cependant, à l’image de ce que nous avons d’ores et déjà remarqué à propos des bœufs, les données relatives aux âges ne permettent pas d’analyser la gestion des troupeaux ni la qualité des viandes consommées dans tous les habitats étudiés. Les données épiphysaires autorisent cependant la comparaison de sept ensembles, dont cinq habitats de hauteur8, un habitat rural9 et une occupation de Bourges “Avaricum”10 (Fig. 9).
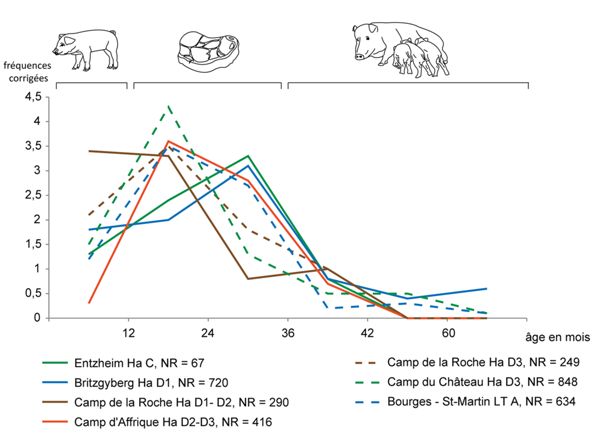
Le taux de reproduction du porc, sa facilité d’engraissement et le fait qu’il ne soit pas utilisé de son vivant pour une quelconque production sont autant d’éléments qui en font l’animal de boucherie par excellence. Il atteint sa maturité pondérale (sans engraissement) aux alentours de deux ans (Vigne 1988), plus rapidement donc que le bœuf ou les caprinés. À ce titre, la présence d’animaux très jeunes dans un ensemble de faune est révélatrice d’une alimentation de qualité, puisque relativement peu dépendante de contraintes économiques. La plupart du temps, les analyses des âges d’abattage des populations porcines révèlent une gestion dite “normale” ou “bouchère”, caractérisée par l’abattage de jeunes animaux de quelques mois, une élimination plus importante d’animaux âgés de près de deux à trois ans et très peu de bêtes plus âgées.
Dans le cas présent, les animaux abattus après trois ans sont très minoritaires, conformément à une gestion raisonnée. Quelques individus sont cependant conservés à un âge avancé, notamment au “Britzgyberg” (Hallstatt D1). Au “Camp de la Roche” (Hallstatt D1-D2), les porcelets âgés de quelques jours à un an sont mieux représentés que partout ailleurs et même davantage que les jeunes bêtes d’un à deux ans. C’est le seul cas de ce genre dont témoignent les données épiphysaires. Ailleurs, les pics d’abattage correspondent aux animaux âgés de un à deux ans11 (qui fournissent une viande de qualité), ou entre deux et trois ans12, âge où les animaux ont atteint leur maximum de rendement carné.
Les données relatives aux autres habitats de hauteur ne peuvent être incluses dans ces analyses par manque de représentativité. Nous pouvons néanmoins noter qu’au “mont Lassois”, dans le secteur du rempart (Hallstatt D2-D3), “la majorité des restes provient de jeunes individus” (Huguet 2011b), laissant ainsi supposer la consommation d’une viande de qualité. En revanche, au “Camp du Châtelet” à Étaules, neuf individus sont âgés de moins d’un an, sept ont été abattus entre un et deux ans, quatre vers trois ans et douze sont âgés de plus de quatre ans (dont deux sont très âgés (Poulain 2003). Ainsi, une partie des animaux fournit une viande de qualité puisqu’elle est abattue avant la maturité pondérale, tandis que l’autre moitié est conservée pour un meilleur rendement boucher ou pour la reproduction. La situation est semblable à Bourges “Hôtel-Dieu” (Hallstatt D2-D3), où l’on constate des parts similaires (aux alentours de 20 %) d’animaux âgés de moins de vingt mois et d’animaux adultes de plus de quatre ans (Augier et al. 2007).
En ce qui concerne les habitats ouverts, nous relevons qu’à Geispolsheim “Kirstenfeld/Bruechel” (Hallstatt D2-D3) les sujets sont abattus en fin de croissance pour la plupart (Méniel 1990), comme à Colmar (Hallstatt D1) où dix individus sur quatorze sont adultes (Poulain 1990). Au “Simlisberg”
(Hallstatt C), en revanche, la plupart des restes osseux correspond à des animaux immatures et les données relatives aux âges dentaires témoignent d’une majorité d’individus juvéniles (Méniel 1991). Enfin, à Entzheim (Hallstatt C) – seul habitat ouvert pour lequel l’analyse de la gestion du cheptel porcin est permise par la quantité de données disponible – le pic d’abattage des porcs se situe entre deux et trois ans, traduisant ici une stratégie d’élevage donnant priorité à la quantité de viande fournie par les animaux, plutôt qu’à sa tendreté13. Dans les autres habitats ouverts, la gestion des cheptels porcins est similaire à celle d’Entzheim, hormis au “Simlisberg” à Morschwiller-le-Bas (Hallstatt C) où la plupart des individus sont immatures, voire juvéniles. Dans ce cas, le fait que les ossements proviennent d’une unique structure isolée ne permet pas de pousser plus avant les interprétations et laisse suspecter une sélection particulière.
En résumé, cette analyse révèle une consommation de viande porcine de qualité fournie par des animaux jeunes dans les habitats de hauteur, ainsi qu’à Bourges “Saint-Martin-des-Champs”. Une nuance semble tout de même pouvoir être apportée dans les cas du “Camp d’Affrique” (Hallstatt D2-D3), du “Camp du Châtelet” (Hallstatt D) et des occupations de Bourges “Hôtel-Dieu” (Hallstatt D2-D3) et “Saint-Martin-des-Champs” (La Tène A). En effet, si la plupart des animaux sont abattus jeunes, les bêtes de boucherie sont tout de même assez abondantes. Cela témoigne d’une volonté de rendement boucher plus forte qui est, peut-être, la conséquence directe d’une démographie plus importante. Dans les habitats ouverts, la stratégie d’élevage semble donner priorité à la quantité de viande fournie par les animaux plutôt qu’à sa tendreté.
Les caprinés, entre ressource carnée de qualité et produits du vivant
La plupart du temps, les caprinés ont un rôle secondaire dans l’alimentation carnée. Bien qu’ils soient fréquents, les masses de restes qui leur sont associées les placent après le bœuf en termes d’apport carné voire, le plus souvent, en troisième position après le porc. Là encore, la faiblesse numérique des données relatives aux âges ne permet pas une étude globale de la gestion des cheptels caprins au Premier âge du Fer, mais conduit cependant à une étude comparative assez précise des habitats “privilégiés”. Les données épiphysaires autorisent, en effet, la comparaison de sept ensembles, dont six correspondent à des habitats de hauteur14 (du Hallstatt D1 au Hallstatt D3) et un dernier associé au complexe proto-urbain de Bourges “Avaricum”15.
La diagnose spécifique des caprinés, lorsqu’elle est permise par l’observation de critères distinctifs ou par l’analyse morphométrique, donne une meilleure représentation des ovins dans la plupart des occupations hallstattiennes (on dénombre en moyenne 3 restes de mouton pour 1 de chèvre). Chez ces deux espèces, la maturité pondérale est atteinte aux alentours de trois ans (Vigne 1988)16 et, à l’instar du bœuf, certaines bêtes sont conservées jusqu’à des âges avancés pour la production laitière, la reproduction, ou encore pour la laine dans le cas des ovins. Les gestions des troupeaux de moutons et de chèvres étant assez différentes du fait de leurs productions respectives, il conviendrait de les différencier. Cela ne peut cependant être fait puisque les diagnoses sont trop rares.
Les âges d’abattage des caprinés révèlent une plus grande diversité dans la gestion des cheptels que pour les bœufs et les porcs (Fig. 10). Sur certains habitats de hauteur, une grande proportion d’agneaux et cabris de lait et une très faible part d’animaux adultes ou réformés traduisent une sélection assez drastique, motivée par une volonté de consommer une viande de qualité provenant de très jeunes animaux. C’est le cas du “Camp de la Roche” (Hallstatt D1-D2) et du “Camp d’Affrique” (Hallstatt D2-D3). Dans ces deux ensembles, la forte représentation des animaux pourrait témoigner de la consommation des bêtes plus âgées dans d’autres secteurs, ou encore d’un approvisionnement en viande depuis l’extérieur.
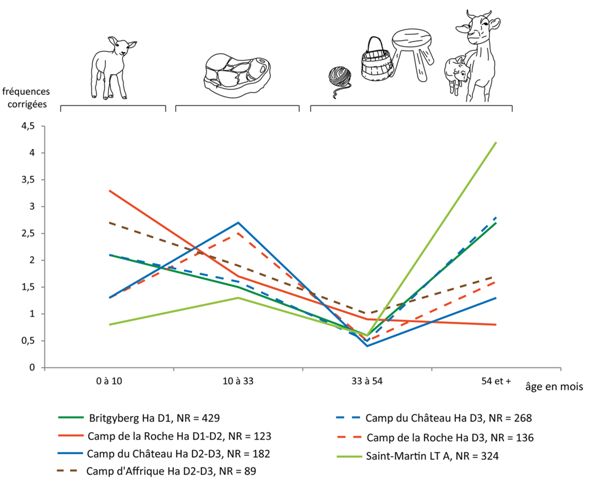
des âges d’abattage des caprinés d’après les données épiphysaires.
Dans d’autres habitats de hauteur, les animaux en bas-âge restent bien représentés, mais une part importante des sujets est abattue après réforme. C’est le cas du “Britzgyberg” (Hallstatt D1) et du “Camp du Château” (durant la seconde phase d’occupation du site au Hallstatt D3). Cette distribution tend à montrer que la recherche d’une viande tendre fournie, une fois encore, par des très jeunes animaux, ne se substitue pas au besoin de s’approvisionner en lait et/ou laine.
Deux autres ensembles traduisent, non plus la sélection d’animaux en bas-âge ni la volonté de développer les productions du vivant, mais le besoin (ou le choix) de produire une viande de qualité en quantité plus importante. Cela se traduit par l’abattage d’animaux plus âgés mais n’ayant pas atteint leur maximum pondéral (de 10 à 33 mois). Il en est ainsi au “Camp du Château” (durant la première phase d’occupation du site au Hallstatt D2-D3) et au “Camp de la Roche” (Hallstatt D3).
Enfin, à Bourges “Saint-Martin-des-Champs” (La Tène A), les animaux sont principalement abattus après réforme, bien que les autres catégories d’âges soient représentées. Sur ce site, la consommation de viande de qualité semble très ponctuelle et l’élevage des caprinés se tourne davantage vers les productions laitière et/ou lainière.
Pour les autres habitats de hauteur considérés dans ce travail, on ne dispose pas de données suffisantes pour les intégrer à cette étude. Nous pouvons cependant mentionner la présence majoritaire de jeunes animaux au “Mont Lassois”, dans le secteur du rempart (Hallstatt D2-D3) (Huguet 2011b), donnant ainsi l’image d’une consommation de viande de bonne qualité. Au “Camp du Châtelet” (Hallstatt D), on dénombre quatre sujets de moins de dix mois, douze individus correspondant à des animaux jeunes fournissant une viande tendre et une cinquantaine de bêtes de réforme, dont trois sont très âgées (Poulain 2003). Il semble donc – à l’image de ce que l’on a pu observer à Bourges “Saint-Martin-des-Champs” – qu’à une faible consommation de viande de qualité viennent s’adjoindre, de manière plus prégnante, des productions laitières et lainières. Au “Buergelen”, les estimations fondées sur les dents traduisent un abattage important d’animaux jeunes (la moitié), âgés de dix-huit mois à trois ans (Méniel 2005), autrement dit de sujets qui sont à l’origine d’une production carnée de qualité. Enfin, à Bourges “Hôtel-Dieu” (Hallstatt D2-D3), “la viande de caprinés est principalement issue d’individus situés autour et après leur optimum de poids, la viande est donc de qualité moyenne à médiocre” (Augier et al. 2007).
Pour les habitats ouverts, on ne dispose que de très rares estimations d’âges. À Geispolsheim “Kirstenfeld/Bruechel” (Ha D2-D3), la majorité des individus est adulte (58 %) (Méniel 1990), traduisant une utilisation développée des animaux de leur vivant. Au “Simlisberg” (Hallstatt C), l’élevage des caprinés semble avoir pour objectif principal la production carnée, la plupart des individus étant abattue jeune (Méniel 1991). Cependant, la faiblesse des données ne permet pas de l’affirmer (NR = 14).
Ainsi, l’élevage des caprinés est susceptible de se tourner préférentiellement vers la consommation de très jeunes animaux – caractéristique d’une situation économique prospère, et peut-être aussi d’un apport extérieur – ou d’animaux n’ayant pas atteint leur maximum pondéral et fournissant ainsi une viande tendre, mais en quantité plus importante. Il peut aussi privilégier les productions du vivant (lait, laine) et ne fournir une production carnée de qualité que de manière ponctuelle voire gérer de manière équilibrée boucherie et productions annexes. Quoi qu’il en soit, le contexte économique et social, la culture ou les goûts – la viande de mouton et de chèvre ne faisant pas l’unanimité encore de nos jours – sont autant d’éléments susceptibles d’être à l’origine de cette pluralité de situations. En ce qui concerne les habitats ouverts, les données sont ici bien trop rares pour déceler d’éventuelles tendances.
Synthèse
De ces analyses nous pouvons tirer cinq conclusions majeures. La première est la différence de statut entre les habitats “privilégiés” et les habitats ruraux, les premiers bénéficiant le plus souvent d’une certaine latitude vis-à-vis des contraintes économiques liées à l’élevage et son rendement en termes d’apport carné. Le second point à souligner est qu’il existe des différences au sein même des habitats privilégiés puisqu’ils ne pratiquent pas les mêmes sélections. Les variations de fréquences des mammifères domestiques sont en partie liées aux conditions environnementales et notamment aux superficies et configurations des plateaux. Cependant, certains d’entre eux ont pu s’affranchir quelque peu de ces contraintes environnementales en se faisant approvisionner par des habitats ruraux. Du reste, les facteurs environnementaux ne suffisent pas à expliquer les écarts de fréquence, qui peuvent aussi être imputés à des différences démographiques, économiques voire culturelles.
À Bourges, les fréquences des mammifères domestiques n’évoluent pas de manière sensible de la fin du VIe s. à la fin du Ve s. a.C., cependant quelques variations semblent exister d’un ensemble à l’autre en fonction du statut des habitants ou des activités pratiquées. Quoi qu’il en soit, la prédominance très nette du porc dans chacun des contextes – observée nulle part ailleurs – nous semble directement liée au statut urbanisé du site.
Dans les habitats ouverts, les variations des fréquences des mammifères domestiques suggèrent une pluralité des formes d’exploitations. Et bien que les données soient lacunaires, les âges d’abattage des animaux semblent traduire la priorité donnée au rendement boucher et aux productions du vivant.
Nous pouvons, enfin, souligner le caractère secondaire de la consommation des viandes canine et chevaline. Dans deux cas cependant, les équidés tiennent une place particulièrement importante dans l’alimentation : au “Buergelen” (faubourg du “Britzgyberg” au Hallstatt D3) et à Wolfgantzen (habitat rural, Haut-Rhin) avec 18 %. L’importance des équidés dans les habitats est donc pour le moins contrastée, à l’image de ce que l’on observe au Second âge du Fer en Gaule (Arbogast et al. 2002). Il n’est pas inconcevable que la consommation de viande chevaline puisse constituer un marqueur social, culturel, ou encore économique.
À ces éléments, et en guise d’ouverture, nous pouvons ajouter la présence de grands bœufs et chevaux dans les habitats de hauteur exclusivement (au “Britzgyberg”, au “Kastelberg” et au “Camp d’Affrique”). Bien que nous ne puissions généraliser ce phénomène, il ne semble pas imprudent d’y voir ici un marqueur social. En effet, pour le bœuf, une taille plus élevée correspond à une force de travail et un poids vif plus importants, ainsi que de plus grandes quantités de lait, de viande et de matières premières. En parallèle, leur entretien nécessite de plus grandes quantités de nourriture et d’eau, ce qui sous-entend la possibilité de fournir cette nourriture de la part des propriétaires. Pour le cheval, une taille élevée est synonyme d’une plus grande rapidité et d’une plus grande force, ce qui peut faire de ces animaux de meilleures montures. Ces grands bœufs et grands chevaux sont donc susceptibles de correspondre à des biens de prestige.
Par ailleurs, une série de quatre dents jugales équines aux dimensions très réduites a été découverte au “Britzgyberg”. Elles correspondent à un très petit cheval, de moins d’un mètre au garrot, dont aucun sujet similaire n’a encore été observé au Premier âge du Fer. L’usure importante des dents témoigne de l’âge très avancé de ce cheval, qui suggère qu’il ne devait plus être en mesure de tirer ou de porter une charge. La conservation de ce cheval après réforme dénote d’un statut généralement octroyé aux animaux “familiers”. En outre, sa taille, son âge et son lieu de découverte (un habitat de hauteur) sont autant d’éléments qui arguent en faveur d’un animal d’apparat.
Pour conclure, la rareté de ces grands animaux et le cas particulier du cheval “nain” du “Britzgyberg” nous amènent à nous questionner sur leur provenance. En effet, le fait qu’ils soient mêlés à des cheptels de tailles différentes, qu’ils ne soient observés que sur des sites au statut privilégié et qu’ils puissent être perçus comme des témoignages d’aisance, sont autant d’éléments qui arguent en faveur d’une importation. Cela reste cependant un point sur lequel il faudra s’attarder à l’avenir en augmentant la représentativité de ces données par de nouveaux apports.17
Bibliographie
- Arbogast, R.-M., Clavel, B., Lepetz, S., Méniel, P. et Yvinec, J.-H. (2002) : Archéologie du cheval : des origines à la période moderne en France, Paris.
- Augier, L., Buchsenschutz, O. et Ralston, I. (2007) : “L’Hôtel-Dieu”, in : Augier et al., éd. 2007, 89‑132.
- Augier, L., éd. (2007) : Un complexe princier de l’âge du Fer. L’habitat du promontoire de Bourges (Cher) (VIe-IVe s. a.C.), Bituriga Monographie 2007-3/RACF Suppl. 32, Bourges-Tours.
- Bonnet, C., Plouin, S. et Lambach, F. (1990) : “Le site protohistorique de Colmar-Sud”, RAE, 41, 1.
- Chaume, B. et Mordant, C., éd. (2011) : Le complexe aristocratique de Vix : nouvelles recherches sur l’habitat, le système de fortification et l’environnement du mont Lassois, Art, Archéologie et Patrimoine, Dijon.
- Desachy, B. (2004) : “Le sériographe EPPM : un outil informatisé de sériation graphique pour tableaux de comptages”, RAP, 3, 1, 39‑56.
- Foucras, S. (2011) : Animaux domestiques et faunes sauvages en territoire arvernes, Archéologie des plantes et des animaux 3, Montagnac.
- Goudemez S. (2018) : Chasse et élevage au Premier âge du Fer dans le nord-est de la France, Éditions Mergoil, Collection Archéologie des Plantes et des Animaux, 6, 263 p.
- Huguet, L. (2011) : “Les vestiges de faune découverts dans le rempart du mont Saint-Marcel”, in : Chaume & Mordant, éd. 2011, 295‑303.
- Lepetz, S. (1996) : L’animal dans la société gallo-romaine de la France du Nord, RAP n° spécial 12, 37.
- Méniel, P. (1990) : “Annexe : La faune du site hallstattien de Geispolsheim ‘Bruechel’ (Bas-Rhin)”, RAECE, 41, 2, 258‑261.
- Méniel, P. (1991) : “Étude des restes animaux du site hallstattien de Morschwiller-le-Bas ‘Simlisberg’ (Haut-Rhin)”, Cahiers de l’APRAA, 7, 115‑120.
- Méniel, P. (2005) : “La faune d’Illfurth”, in : Roth-Zehner, éd. 2005.
- Nicolardot, J.-P., éd. (2003) : L’habitat fortifié pré- et protohistorique en Côte d’Or. Les camps de Myard à Vitteaux et du Châtelet d’Etaules dans le contexte archéologique régional (du Ve millénaire au IVe siècle avant J.-C.), RAE Suppl. 19, Dijon.
- Poulain, T. (1990) : “La faune”, RAE, 41, 1, 39‑45.
- Poulain, T. (2003) : “Châtelet d’Étaules (faune mise au jour avant 1996)”, in : Nicolardot, éd. 2003, 223‑234.
- Roth-Zehner, M., éd. (2005) : Illfurth (68) : “Buergelen”, lotissement “Les hauts de Buergelen”, Rapport de fouilles préventives, ANTEA inédit.
- Vigne, J.-D. (1988) : Les mammifères post-glaciaires de Corse : étude archéozoologique, Gallia Préhistoire Suppl. 26, Paris
Notes
- Cerf, lièvre, sanglier, chevreuil, aurochs, castor, renard, chat, loutre, martre ou fouine, loup, blaireau.
- Perdrix sp., pigeon ramier, milan noir, milan royal, cygne, grue, canard sp., oie, grand tétra, engoulevent.
- Les masses des restes ne sont pas renseignées dans les ensembles de Colmar, de Besançon et du “Camp du Châtelet” à Etaules.
- Besançon “Saint-Paul” (Doubs).
- “Collège Littré” (Hallstatt D2-D3) et Saint-Martin-des-Champs (La Tène A).
- “Britzgyberg” (Hallstatt D1), “Camp de la Roche” (Hallstatt D1-D2), “Camp d’Affrique” (Hallstatt D2-D3 et Hallstatt D3-La Tène A), “Camp du Château (Hallstatt D2-D3 et Hallstatt D3).
- Les âges ne sont pas précisés par l’auteur.
- “Britzgyberg” (Hallstatt D1), “Camp de la Roche” (Hallstatt D1-D2 et Hallstatt D3), “Camp d’Affrique” (Hallstatt D2-D3), “Camp du Château (Hallstatt D3).
- Entzheim (Hallstatt C).
- “Saint-Martin-des-Champs” (La Tène A).
- Les pics d’abattage se situent entre un et deux ans au “Camp de la Roche” (Hallstatt D1-D2 et Hallstatt D3), au “Camp d’Affrique” (Hallstatt D2-D3), au “Camp du Château” (Hallstatt D3) et à Bourges “Saint-Martin-des-Champs (La Tène A).
- Les pics d’abattage se situent entre deux et trois ans au “Britzgyberg (Hallstatt D1) et à Entzheim (Hallstatt C).
- Là encore, il est probable que la conservation différentielle soit à l’origine de la sous-représentation des animaux juvéniles. Ces résultats doivent donc être considérés avec la prudence nécessaire.
- Le “Britzgyberg” (Hallstatt D1), le “Camp de la Roche” (Hallstatt D1-D2 et Hallstatt D3), le “Camp du Château” (Hallstatt D2-D3 et Hallstatt D3), le “Camp d’Affrique” (Hallstatt D2-D3).
- “Saint-Martin-des-Champs” (La Tène A).
- Dans le cadre de cette étude comparative, la limite de maturité pondérale est fixée à trente-trois mois, mais cela ne diminue en rien la représentativité des données.
- Depuis la rédaction de cet article en 2016, l’intégralité des recherches menées dans le cadre de ce doctorat a été publiée (Goudemez 2018).
